

Dal 2 al 4 dicembre 2010 si terrà ad Hammamet il 4th Mediterranean Symposium on Marine Vegetation, un workshop dedicato dove si discuterà anche di fanerogame marine e della loro indiscutibile importanza nell’ambiente marino. Alcune associazioni, come la World Seagrass Association, promuovono da qualche anno lo studio e la ricerca sulle fanerogame, anche se purtroppo, ultimamente, l’attività dell'associazione è ridotta e il sito non viene aggiornato da tempo.
Le fanerogame (faneròs = evidente; gàmos = nozze), chiamate impropriamente alghe, sono in realtà delle vere e proprie piante a cormo, con radici, fusti, foglie e fiori, quindi sono molto diverse dalle alghe vere e proprie. Sono delle angiosperme monocoltiledini perfettamente adattate alla vita acquatica, tanto da non poter sopravvivere in ambienti diversi perchè prive di qualunque sistema atto ad accumulare o prevenire la perdita dell’acqua stessa. I pollini inoltre sono privi dei caratteristici rivestimenti che permettono ai pollini delle piante terrestri di viaggiare nell’aria.
Ma quali sono le differenze tra fanerogame e le alghe?
Le alghe sono organismi meno evoluti, mono e pluricellulari, ma mai organizzate in tessuti veri e propri. L’intera struttura si presenta indifferenziata e svolge la stessa funzione, con l’esclusione degli sporangi e dei gametangi che producono rispettivamente spore e gameti. Le fanerogame invece, come accennato sopra, sono in grado di produrre organi differenziati e con funzioni differenziate come radici, fusti (rizomi), foglie, fiori, frutti e semi. La riproduzione avviene per stolonizzazione e grazie a fenomeni gamici che terminano con la produzione del seme.
L'ORIGINE DELLE FANEROGAME
Si ritiene che le prime fanerogame marine si siano diffuse nei mari durante il medio e tardo Cretaceo, ovvero circa 100 milioni di anni fa. La teoria di Arber afferma che le fanerogame si siano originate da piante di acqua dolce che hanno poi affrontato un improvviso aumento di salinità e si siano diffuse progressivamente dalle acque dolci, a quelle salmastre per poi arrivare al mare. La teoria di Den Hartog afferma invece che le fanerogame si siano originate più semplicemente da piante terrestri alotolleranti, ovvero da specie che vivevano nelle zone intertidali e adattatesi a fasi periodiche di emersione e immersione nell’acqua di mare.
Nelle acque dolci esistono moltissime specie di fanerogame, e poche nelle acque marine. Dal confronto tra il numero di specie nei due ambienti, Arber dedusse che proprio gli ambienti di acqua dolce fossero i luoghi di origine delle fanerogame. Ma a sostegno dell'ipotesi di Den Hartog vi sono delle prove più convincenti. Per esempio nelle mangrovie (che sono piante alotolleranti), le basi fogliari racchiudono le foglie più giovani per proteggerle dalla salinità. Inoltre i tessuti delle fanerogame di acque dolci sono assai meno complessi di quelli delle fanerogame marine. I fusti di queste ultime rimangono attaccati alla pianta madre (viviparità vegetale). Purtroppo non esistono molte testimonianze fossili, solo del tardo cretaceo esistono fossili relativamente numerosi dei generi Posidonia (come P. cretacea e P. parisientis, conosciuta anche come P. perforata oppure Caulinites parisientis. Quest'ultima proviene da sedimenti Eocenici del Paris Basin) e Thalassocharis. Le analisi di filogenesi molecolare hanno evidenziato invece l'origine polifiletica delle fanerogame, ma su questo punto occorre attendere ulteriori studi.
Riguardo alla zona di origine, si pensa che l'area Indonesiana, del Borneo e della Nuova Guinea, che vanta il maggior numero di specie, sia stato il sito di origine delle fanerogame. Poi si sarebbero diffuse nel resto del mondo (Ipotesi del centro di origine). L'ipotesi della vicarianza afferma invece che la distribuzione sia stata abbastanza uniforme lungo le coste dell'Africa orientale e dell’Indonesia. Le fanerogame, escluse le coste antartiche, sono diffuse ovunque.

Prateria a fanerogame. Fotografia di Castronuovo M. Nicola
MORFOLOGIA DELLE FANEROGAME
Le fanerogame sono piante modulari composte da unità dette ramet. Le ramet secondo la definizione di T. Smith sono membri individuali, ovvero cloni, di una pianta modulare. Nelle piante stolonifere come le fanerogame, i cloni si espandono lateralmente con fusti e successivamente con nuove radici che poi si separano fisiologicamente dalla pianta madre. Per esempio, in Posidonia oceanica e in altre fanerogame, è il rizoma orizzontale (o rizoma plagiotropo), un fusto modificato e strisciante che penetra nel substrato, sul quale, in prossimità dei nodi, si formano le nuove radici e successivamente i nuovi stoloni. Come si vede nella figura sottostante i rizomi sono dimorfici, ossia orizzontali ma anche verticali (o rizomi ortotropi), e quelli più corti sono in genere indicati con il termine stem. La funzione del rizoma plagiotropo è quella quindi di ancorare la pianta al substrato, e permetterne l’avanzamento in senso orizzontale e formare le cosiddette praterie.

Il rizoma ortotropo permette invece la crescita della pianta in senso verticale e superare così eventuali condizioni avverse come quelle dovute al seppellimento da parte del sedimento.
Entrambe le tipologie di rizomi possono passare da una forma all’altra. Un rizoma ortotropo può divenire plagiotropo quando cessa la crescita verticale, oppure un rizoma plagiotropo può divenire ortotropo con la morte del meristema apicale. I rizomi nelle diverse specie si distinguono anche per la differente elasticità, quelli verticali sono flessibili e adattabili ai movimenti dell’acqua, quelli orizzontali più robusti e tendenti alla lignificazione. Più in generale i rizomi costituiscono lo strato ipogeo della pianta, molto stabile nel tempo, mentre le foglie costituiscono lo strato epigeo, molto variabile in funzione delle condizioni ambientali e stagionali.
L’apice vegetativo, costituito da 6 o 7 giovani foglie è situato proprio nella parte superiore del rizoma. Le foglie più giovani sono interne e protette dalle più vecchie che si presentano più lunghe e robuste. La funzione delle foglie è quella fotosintetica ma anche quella di assorbire nutrienti dall’ambiente acquatico. Esse hanno un parenchima specializzato detto aerenchima o tessuto aerifero, o anche lacune aerifere, caratterizzato da tanti piccoli spazi pieni di aria che permettono il galleggiamento delle foglie e gli scambi gassosi con l’esterno. Se la foglia viene danneggiata, l’ingresso dell’acqua e il relativo allagamento della foglia è impedito dai setti che caratterizzano le lacune. Le cellule epidermiche delle foglie contengono cloroplasti (a differenza delle monocotiledini terrestri che hanno i cloroplasti nel mesofillo), numerosissimi mitocondri e sono caratterizzate da un plasmalemma molto invaginato allo scopo di aumentare la superficie di scambio. La cuticola fogliare è molto sottile e stomi e le cellule di guardia sono assenti. Le venature delle foglie sono generalmente parallele. Le foglie possono essere molto lunghe come nel caso di Halophyla e di Posidonia, che hanno o possono avere foglie lunghe più di un metro, oppure corte, solo 30 – 40 cm come in Zoostera nana. I vasi sono sottili e non lignificano come nelle piante terrestri. Le foglie si accrescono alla base, quindi l’apice è la parte più vecchia della foglia stessa e per primo andrà incontro a fenomeni di senescenza. Quando la foglia si stacca, alla base rimangono delle scaglie (lipule) che andranno incontro a lignificazione e il rizoma risulterà ricoperto da esse a guisa di manicotto. Le scaglie permettono di risalire all’età della pianta e con una certa approssimazione anche a quella della prateria. Il loro spessore viene misurato in mm e permette di risalire all’andamento della crescita della pianta.
Altri adattamenti che le fanerogame hanno dovuto escogitare riguardano le carenze di ossigeno, infatti spesso i sedimenti ove si accrescono sono anossici (privi o scarsi di ossigeno), e quindi esse accumulano ossigeno proprio nelle radici, ove giunge attraverso i rizomi e il parenchima aerifero delle foglie, quest’ultime sono la sede della fotosintesi e quindi la fonte di ossigeno per l'intera pianta.
La riproduzione delle fanerogame marine può essere assessuata o sessuata. Nel primo caso si parla di stolonizzazione, gia accennata. Nuovi rizomi si originano da quelli preesistenti dando origine a dei cloni che si espandono lateralmente originando una struttura intricata ove diviene difficile distinguere le singole unità. La riproduzione sessuale invece avviene grazie alla produzione di fiori, frutti e semi. Meno del 10% degli individui di una prateria a Posidonia utilizza questa modalità di riproduzione, mentre nel caso dei generi Halophyla e della specie Enhalus acoroides la fioritura è fequente. I fiori si originano dai nodi dei rizomi e sono piccoli e difficili da vedere per un occhio inesperto. Alcune specie comunque hanno fiori appariscenti, altre ancora sviluppano fiori solitari apicali. Il periodo della fioritura è diverso per ogni specie. In Mediterraneo Zostera marina fiorisce da aprile a giugno, Cymodocea nodosa a maggio, Posidonia oceanica infine fiorisce a settembre - ottobre.
Nelle Posidoniaceae i fiori sono actinomorfi (a simmetria raggiata) ed ermafroditi. Sono presenti tre stami e un solo carpello, elissoide e irregolare. Lo stigma è sessile e ornato.
L’impollinazione può essere subaerea, efidrofila e idrofila. Nel caso di fanerogame di acque dolci il fiore spunta fuori dall’acqua per cui l’impollinazione è subaerea. Negli altri casi il polline è idrorepellente e gelatinoso, e viene disperso in acqua (impollinazione idrofila) o sulla sua superficie (impollinazione efidrofila). In mare solo la specie Enhalus acoroides si affida all’ impollinazione subaerea. Il polline del genere Posidonia è filiforme e le specie sono prive di un sistema per impedire l’autoimpollinazione.
I frutti delle fanerogame possono essere drupe, acheni, capsule ecc. Le drupe di P. oceanica galleggiano e consentono alla specie di colonizzare nuovi ambienti, mentre gli acheni di Zostera marina rimangono in situ senza allontanarsi molto dal sito di origine. Alcune specie come Talassodendron sono vivipare ovvero i semi si sviluppano prima di lasciare la pianta madre. In alcune specie di Posidonia l’ipocotile può rimanere attaccato anche 18 mesi, mentre la vitalità del seme non supera gli otto mesi (Hocking et al., 1980).
ECOLOGIA DEI SISTEMI A FANEROGAME

Come accennato, le fanerogame sono quasi tutte fotofile e colonizzano prevalentemente il piano infralitorale, ove vi è luce a sufficienza. Tuttavia alcune specie in acque particolarmente trasparenti colonizzano fondali sino a 50 metri di profondità. Più in generale vale la regola secondo la quale le fanerogame crescono in zone ove vi sia almeno il 10 – 20 % della luce incidente superficiale. Quindi sono importanti fattori limitanti la trasparenza delle acque, la presenza di acidi umici, la presenza di foci di fiumi e corsi d’acqua, e non da ultimo, le attività umane, che inquinano pesantemente ogni ambiente. Il valore del 20% può sembrare alto, ma le fanerogame necessitano di alti tassi fotosintetici per produrre ossigeno e veicolarlo ai rizomi e alle radici, spesso impiantate in ambienti anossici e privi quindi di ossigeno. Alcune fanerogame del genere Halphyla sono state trovate sino a 58 metri di profondità, in acque molto trasparenti della Grande Barriera Corallina Australiana. Le fanerogame possono anche manifestare fotoinibizione in zone troppo illuminate. Ricordiamo che in acque molto trasparenti la luce giunge con valori accettabili per le fanerogame anche a maggiori profondità, ma poi subentra un altro fattore limitante, la pressione idrostatica. È noto per esempio che Halodule uninervis non cresce a grandi profondità non per mancanza di luce ma per l’eccessiva pressione idrostatica.
Nelle zone intertidali i limiti delle escursioni di marea influenzano la distribuzione di numerose specie, alcuni studi hanno evidenziato fotoinibizione, diseccamento e danni da raggi UV, durante la fase di bassa marea. La salinità in alcune specie, come per esempio Zostera novazelandica influenza l’abbondanza della fioritura e quindi la riproduzione sessuale. Anche la germinazione dei semi sembra essere influenzata dalla salinità.
Anche la tipologia del substrato è importante. Per esempio Cymodocea nodosa si impianta sia su substrati puliti che putridi, mentre Posidonia oceanica è molto meno tollerante e colonizza solamente substrati puliti. Sempre P. oceanica è in grado di trattenere gran quantità di sedimento e prevenire così efficacemente l’erosione, e costruire le cosidette mattes, per questo ospita ricche comunità animali. L’intera prateria di P.oceanica potrebbe benissimo essere considerato un unico organismo bioindicatore.
L’accrescimento delle foglie di P. oceanica è massimo in primavera e minimo in estate, e la densità dei ciuffi varia da meno di 50 a oltre 700 per m2. Se è minore di 50 significa che la prateria è in regressione, sopra i 100 significa che la prateria è in buona salute. P. oceanica ma anche altre fanerogame, per evitare la predazione hanno escogitato delle difese chimiche. Le foglie sono ricche di tannini (proantocianine), ma anche di acidi fenolici e così non vengono brucate da organismi predatori.
Le praterie di fanerogame, in particolare quelle a P.oceanica sono importantissime dal punto di vista ecologico. Nelle nuove foglie molto presto si sviluppa un fauling, ovvero un biofilm di piccoli organismi epifiti (epibionti vegetali) che si distribuiscono con successioni spaziali ben precise sulle foglie. Moltissimi briozoi e alghe si distribuiscono all’apice, che è la zona più ricca di luce. Tra le alghe ricordiamo Ceramium (un'alga rossa), Polysiphonia (altra alga rossa), Castagnea (un'alga bruna), Dictyota (altra alga bruna) e Giraudia. L'insieme di queste associazioni è detto Posidonietum. Tra gli invertebrati fotofili ricordiamo Sertularia perpusilla (Idrozoo), Electra posidoniae (Briozoo), Fenestrulina joannea (Briozoo).
Alla base delle fanerogame sono presenti organismi sciafili che non necessitano di molta luce, come idozoi, briozoi (Schizobrachiella sanguine), ascidie (Halocynthia papillosa e Microcosum sulcatus) e spugne (Sycon ciliatum, Dyssidea fragilis). Tra esse sono presenti poi molluschi di varie specie, nematodi, dominanti in autunno, e copepodi arpacticoidi la cui crescita è sincronizzata con gli epifiti delle fanerogame. L’intricato reticolo dei rizomi è popolato da diverse specie di policheti (Lysidice ninetta, Lysidice collaris e Nematonereis unicornis), detti borers, che scavano lunghissime gallerie e capaci di attaccare anche i rizomi, di crosatcei decapodi anch'essi scavatori di gallerie e infine da echinodermi.
Altri organismi che vivono tra le fanerogame si nutrono degli epifiti e delle foglie, per esempio Sarpa salpa (Velimirov, 1984), è in grado di utilizzare le foglie di P. oceanica. Altri potenziali consumatori di P. oceanica sono l’isopode Idotea baltica (Lorenti and Fresi, 1983), l’echinoide Paracentrotus lividus (Zupo and Fresi, 1984), il polichete Platynereis dumerilli (Mazzella et al., 1995), e infine anche la tartaruga Chelonia mydas (Hays et al., 2002). L’importanza di questi macroconsumatori è tale da garantire un controllo delle praterie, un tempo comunque molto più estese e ora in regressione a causa delle attività antropiche. In Australia, gli Isopodi consumano direttamente le foglie delle Posidonie, a differenza di quelli del bacino del Mediterraneo che consumano gli epifiti.
Isopodi, policheti, amfipodi, decapodi e molluschi sono importanti poiché trasferiscono parte della produzione primaria ai livelli superiori della catena trofica. A proposito della produzione primaria, nelle praterie a P. oceanica la produzione va da 68 a 147 gr. Carbonio/m2 all'anno (foglie) e da 8 a 18 gr. Carbonio/m2 all'anno (rizomi). Le specie australiane di Posidonia risultano maggiormente produttive, con valori che arrivano a 1.100 gr. Carbonio/m2 all'anno. Di tale produzione, un 3-10% è utilizzata dagli erbivori, il 23-34% entra nella catena del detrito (ovvero è utilizzato dagli organismi detrivori) e infine il 27-35% è esportato.
Circa 50 specie di pesci sono associati direttamente o indirettamente alle praterie di P. oceanica. Il 56% di queste sono residenti, il 22% transienti e il 22% occasionali. Presenti Scorpenidi, Serranidi, Labridi e Centracantidi. I rizomi ospitano invece fitocenosi sciafile, che ospitano a loro volta briozoi epifitici e altri piccoli organismi. Le specie che crescono alla base dei rizomi sono oltre 70 (Piazzi et al., 2002). Le comunità delle praterie di P. oceanica variano comunque in relazione alla profondità delle praterie. Quelle descritte sono comunità che si riscontrano principalmente nei primi metri della colonna d’acqua, ma P.oceanica può crescere sino a 50 m. di profondità. Le comunità profonde sono meno conosciute. Alcuni studi hanno evidenziato che le comunità a Decapodi sono maggiormente numerose in praterie posizionate a 6 – 10 metri di profondità. In alcune stazioni a 6 metri, sono state censite estese comunità a Cestopagurus timidus, Calcinus ornatus e Pagurus chevreuxi. Si tratta di specie abili a salire sulle foglie di P. oceanica e a nutrirsi degli epifiti. Scendendo con la profondità, le stesse comunità pian piano si riducono. Sono state proposte tre suddivisioni per le comunità a Decapodi:
- comunità superficiali (shallow water groups) < 6 metri
- comunità intermedie (intermediate group) 6 – 11 metri.
- comunità profonde (deep-water group) > 21 metri.
Poco studiata anche la catena del detrito associata alla enorme biomassa prodotta dalle fanerogame, a cui sono associati molti organismi decompositori come protozoi, funghi e batteri. L’ammasso di foglie, che prende il nome di banquette è anche rifugio di moltissime specie di organismi marini. Tra i parassiti di P. oceanica ricordiamo il pirenomicete Hallothia posidoniae e Pontoporeia bilurbinata. Tra i mesofilli delle specie australi P. ostenfeldii e P. australis è stata descritta rispettivamente la presenza degli isopodi Lynseia annae e L. dianea.
DISTRIBUZIONE
Le specie di fanerogame marine sono pochissime rispetto alle monocotiledini terrestri. Sono note circa 60 specie a fronte di oltre 300.000 specie di angiosperme conosciute. Le fanerogame sono presenti nelle regioni temperate e tropicali. Riguardo alla distribuzione, le regioni temperate boreali sono dominate dal genere Zostera, mentre quelle australi dal genere Posidonia, in entrambi gli emisferi sono comunque presenti i duei generi. Nella figura sottostante sono riportati i mari del globo suddivisi per regioni, che sono 10.

Regioni di distribuzione delle fanerogame marine. Per le specie delle singole regioni vedere qui. Fonte: Global seagrass research methods di Frederick T. Short, Robert G. Coles - Elsevier Science 2001.
Ill genere Zostera è maggiormente diffuso, soprattutto nell’emisfero nord, mentre ai due lati del Pacifico è presente il genere Phyllospadis. Specie congeneri vivono nelle regioni temeprate a nord e a sud del globo. E’ il caso di Posidonia, presente in Mediterraneo, con la specie endemica P. oceanica e in Australia con le specie P. australis, P.angustifolia , P. sinuosa e P. ostenfeldii. Sul genere Posidonia sono disponibili anche diversi studi di filogenesi molecolare e hanno mostrato una divergenza tra P. oceanica e le specie australiane, in almeno due regioni del DNA, mentre gli stessi studi hanno evidenziato che le specie P. ostenfeldii , P. coriacea, P. denhartogii, P. kirkmanii, P. robertsoniae sono tutte appartenenti al P. ostenfeldii complex, ovvero sono un’unica specie.
LE FANEROGAME DEL MEDITERRANEO
Nel bacino del Mediterraneo sono presenti diverse specie di fanerogame, quali la gia menzionata Posidonia oceanica, Cymodocea nodosa, Zostera marina, Nanozostera noltii (o Zostera noltii) e Halophila stipulacea. Lungo le coste egiziane del Mediterraneo è stata trovata anche Halodule uninervis. Negli ambienti salmastri e lagunari sono presenti Ruppia maritima e Ruppia cirrhosa.
P. oceanica ((L.) Delile, 1813) è la più conosciuta e anche la più facile da riconoscere, è presente in tutto il bacino del Mediterraneo sia su fondali sabbiosi ma anche su rocce e coralligeno. Presente sino a 50 metri di profondità, ma più comunemente la si trova sino a 20 – 30 metri di profondità. La si riconosce per il caratteristico rizoma con scaglie (coltre di resti di foglie morte), per le foglie molto lunghe, anche 1 metro, e perchè forma intricate ed estese praterie un tempo molto diffuse lungo i litorali delle acque italiane. E’ una pianta monoica, ovvero i fiori hanno stami e carpelli. L'accrescimento verticale dei rizomi di P.oceanica porta alla formazione di una struttura chiamata matte, costituita da un intreccio di rizomi morti e radici tra i quali resta intrappolato il sedimento. Solo la parte superiore di queste strutture è formata da piante vive. La formazione delle mattes dipende in massima parte dai ritmi di sedimentazione; un'alta velocità di sedimentazione può portare ad un eccessivo insabbiamento dei rizomi e quindi al loro soffocamento; al contrario, una sedimentazione troppo lenta può portare allo scalzamento dei rizomi ed alla regressione della prateria. Poiché la velocità di decomposizione dei rizomi è molto lenta essi possono rimanere all'interno della matte anche per millenni. La matte ha un ritmo di crescita molto lento: il suo accrescimento è stato stimato in circa 1 m al secolo.
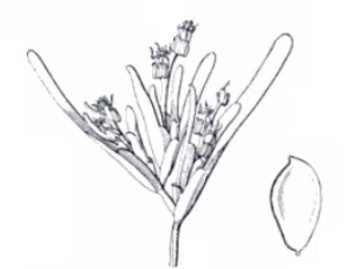
Infloresenza e frutto di P. oceanica (Fonte: R. M. T. Dahlgren, H. T. Clifford, Peter F. Yeo - Families of the Monocotyledons: Structure, Evolution, and Taxonomy – Springer.
Fiorisce, seppur raramente, tra settembre e ottobre, con fiori poco vistosi ma di un bel rosso nella parte interna e verdi esternamente, mentre fruttifica verso marzo - aprile. I fiori sono ermafroditi. I frutti sono detti “olive di mare”, si presentano con un pericarpo poroso e ricco di sostanze oleose che ne consentono il galleggiamento e quindi la dispersione ad opera delle correnti e/o del vento anche in luoghi moto lontani. Poco dopo il pericarpo si rompe, e il seme uscendo cade sul fondale ove, nel caso di condizioni idonee, può germinare e dar vita d una nuova prateria. La fioritura di P.oceanica dipende da fattori ambientali o esogeni come l’irradiazione solare e la temperatura dell’ambiente, e da fattori interni o endogeni come l’età della pianta. Anche il numero e la densità dei germogli influenzano la fioritura. In autunno P. oceanica perde le foglie più vecchie, che le mareggiate vanno ad accumulare lungo le coste sottoforma di cuscini che possono raggiungere anche spessori notevoli. Spesso accanto a questi cuscini si trovano le famose palline di posidonia o egagropili (o posidonia balls in lingua anglosassone) piccoli agglomerati sferici, ovali e a volte irregolari, di colore marrone formati dai residui fibrosi della Posidonia e intrecciati con tanta cura dall’azione delle onde. Sono resistentissimi e possono rimanere invariati per anni.

Egagropile (o posidonia ball in lingua anglosassone). Fotografia di Castronuovo M. Nicola
Cymodocea nodosa è la seconda specie per diffusione in Mediterraneo dopo P. oceanica. Preferisce substarti sabbiosi e siti riparati dal moto ondoso. Vegeta abbondante nei primi metri di profondità, ma la si può trovare anche a 40 metri. A differenza di P.oceanica, che presenta praterie geneticamente poco distinte, quelle di C. nodosa sono invece geneticamente diversificate. Nelle zone di regressione delle praterie a P. oceanica o su matte morte, C. nodosa può insediarvisi insieme a N. noltii. La specie è dioica. Presso la foce dell'Isonzo è nota come "baro canela" che significa "alga cannuccia" perchè i rizomi rossastri ricordano la cannuccia palustre. Fiorisce a primavera e fruttifica in estate.
Zostera marina è diffusa solo nel nord Adriatico e preferisce luoghi ove vi siano apporti di acque dolci. Non è diffusa altrove quindi la sua distribuzione è limitata. Si tratta di una specie relitta. Presente, con popolazioni puntiformi, anche lungo le coste marchigiane. La specie è monoica. In laguna la sua diffusione è limitata dalla salinità, quando raggiunge un valore simile a quello dell'acqua marina entra in azione un fungo parassita che ne arresta la crescita. E infatti nell'inverno del 1929 estese popolazioni marine sono state decimate e fortunatamente sono state risparmiate le popolazioni lagunari perchè la salinità risulta troppo bassa, attorno al 20 per mille. Per cui la pianta è poco diffusda in mare, a dispetto del nome, e molto diffusa in zone salmastre. E' pascolata anche dall'avifauna delle zone salmastre, poiché cresce gia a pochi decimetri di profondità. La fioritura avviene a primavera. Le infiorescenze sono delle spighe chiuse da due lembi di foglie sovrapposte, da cui fuoriescono solo gli stimmi.
Nanozostera noltii un tempo diffusa in tutto il Mediterraneo, predilige substrati a granulometria fine e ove sono in atto processi riduttivi, in zone salmastre o di fronte alle foci dei fiumi. La specie è monoica. La pianta sopporta molto bene l'emersione e la si trova quindi anche nelle zone intertidali. Nella laguna veneta la specie è in rarefazione e negli ultimi anni sta scomparendo a tassi molto elevati.
Halophila stipulacea è una specie lessepsiana, comune nel mar Rosso e insediata lungo le coste Mediterranee sin dal secolo scorso nella parte est del bacino, e sin dalla seconda metà degli anni '80 nella parte ovest del Mediterraneo. Forma praterie molto dense, e presenta un maggior numero di fiori maschili rispetto ai fiori femminili. Presente lungo le coste calabresi e siciliane, fiorisce e fruttifica tutti gli anni.
CLASSIFICAZIONE DELLE PRATERIE A POSIDONIA

Prateria fotografata nelle acque antistanti Selinunte. Fotografia di Castronuovo M. Nicola
Lo studio delle praterie prevede l'analisi di molti aspetti legati alla tipologia del substarto. Sono da considerare la litologia del sedimento, il profilo costiero e la batimetria. Sulla base delle diverse caratteristiche possiamo distinguere diversi tipi di prateria:
-praterie su roccia e coralligeno, formate solo da P. oceanica;
-praterie su sabbia, che possono essere formate da tutte le fanerogame del Mediteraneo;
-praterie su strutture biogeniche, formate dall'intrico di rizomi, radici ecc.; quelle di P. oceanica prendono il nome di matte, quelle di C. nodosa e N. noltii si chiamano turf;
Inoltre le praterie possono essere pure o monospecifiche, oppure miste o plurispecifiche. Le prime sono formate esclusivamente da una specie e sono più comuni a batimetrie maggiori, le seconde sono formate da più specie e sono più comuni a basse profondità. La classificazione delle praterie non termina qui, esistono classificazioni più tecniche e proprie, anche per i termini utilizzati, degli specialisti. Non è raro incontrare sui testi universitari termini come herbier tigrè che indica la prateria tigrata, ovvero fatta di cordoni di faenoragame che si alterano a cordoni di sabbia e paralleli alla linea di costa, oppure ancora herbier de colline, herbier de plain. In questa sede ci preme ricordare le récif barrière o praterie di barriera, presenti esclusivamente in bassi fondali e in zone molto riparate. Si formano delle vere barriere che nel tempo daranno vita a una piccola laguna interna. La barriera infatti si accresce man mano che si deposita sedimento e con la crescita della pianta. Il fronte esterno della barriera si evolve accrescendosi verso il largo, quello interno verso la linea di costa si evolve con la sostituzione di P. oceanica con C. nodosa e N. noltii.

Un récif barrière o praterie di barriera. Fotografia di Castronuovo M. Nicola
BIBLIOGRAFIA
- Borg J – Specie richness and abundance of decapods crustaceans associate with a Maltese Posidonia oceanica. University of Malta.
- Frederick T. Short, Robert G. Coles - Global seagrass research methods - Elsevier Science 2001
- Hocking et al. – Nutrient accumulation in the fruits of two species of seagrasses, P. australis e P. sinuosa. Ann. Bot. 45: 149-165. 1980
- Mattia Barsanti, Ivana Delbono, Ornella Ferretti, Andrea Peirano, Carlo Nike Bianchi and Carla Morri - Measuring change of Mediterranean coastal biodiversity: diachronic mapping of the meadow of the seagrass Cymodocea nodosa (Ucria) Ascherson in the Gulf of Tigullio (Ligurian Sea, NW Mediterranean). Hydrobiologia Volume 580, N.1 / Aprile, 2007 Springer Netherlands
- Marbà N, & Duarte C M -Coupling of seagrass (Cymodocea nodosa) patch dynamics to subaqueous dune migration. Journal of Ecology, 83: 381-389
ARTICOLI CORRELATI
- LE ALOFITE
- LA VEGETAZIONE DEGLI AMBIENTI DUNALI
- LE PIANTE PSAMMOFILE
- Posidonia oceanica, CROCE E DELIZIA DELLE NOSTRE COSTE
- LA FLORA DEGLI STAGNI
Questo articolo è protetto da Copyright © e non può essere riprodotto e diffuso tramite nessun mezzo elettronico o cartaceo senza esplicita autorizzazione scritta da parte dello staff di BiologiaMarina.eu.
Ideazione: Pierfederici Giovanni - Progetto: Pierfederici Giovanni, Castronuovo Motta Nicola.
Prima Pubblicazione 31 Lug 2006 - Le immagini dei Collaboratori detentori del Copyright © sono riproducibili solo dietro specifica autorizzazione.
Si dichiara, ai sensi della legge del 7 Marzo 2001 n. 62 che questo sito non rientra nella categoria di "Informazione periodica" in quanto viene aggiornato ad intervalli non regolari
XHTML 1.0 Transitional – CSS
