
TRILOBITI - 1° parte
Dopo aver descritto alcuni giacimenti interessanti per il loro contenuto paleontologico, con questo breve e certamente non esaustivo articolo, vogliamo parlarvi di alcuni organismi marini, assenti da milioni d'anni nei mari del nostro pianeta, ma molto comuni e diffusi nei mari della preistoria: i trilobiti.
Se potessimo tornare indietro nel tempo, ed immergerci nei mari di 450 milioni di anni fa, probabilmente vedremmo brulicare centinaia di trilobiti, alcuni grandi e corazzati, altri piccolissimi e indifesi.
I trilobiti appartengono al phylum degli Artropodi; quest'ultimi sono animali metamerici (con corpo suddiviso in segmenti), con un esoscheletro che può essere chitinoso, chitinoso-calcareo, chitinoso-fosfatico o addirittura interamente calcareo. Tale esoscheletro, di origine epidermica, negli attuali artropodi ricopre interamente il corpo e presenta segmenti articolati che permettono all'animale di muoversi.
Nei trilobiti invece, pur essendo questi i primi artropodi a secernere un esoscheletro calcitico, le zampe risultavano libere o presentavano limitati processi di calcificazione, per questo motivo trovare fossili completi dei trilobiti è davvero difficile.
L'attuale sistematica colloca i trilobiti nel primitivo gruppo degli Archeocrustacea, tuttavia una loro precisa collocazione, come riporta anche Wikipedia, è ancora incerta
MORFOLOGIA
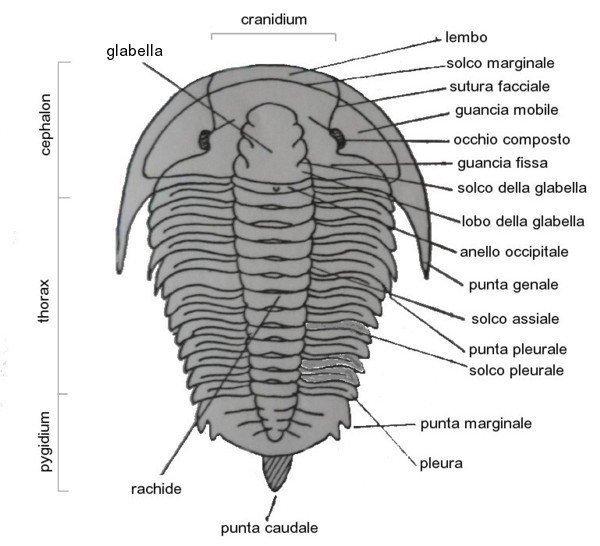
I trilobiti devono il loro nome al fatto che il loro corpo può essere ipoteticamente diviso longitudinalmente e trasversalmente in tre parti, o lobi; in senso longitudinale possiamo osservare uno scudo cefalico (cephalon) costituito da tergiti (segmenti) fusi, un torace (thorax), flessibile e suddiviso in tergiti (fino ad un massimo di 30) e uno scudo caudale (pygidium) formato da una serie di tergiti fusi insieme; trasversalmente possiamo osservare due pleure laterali (1DX, 1SX) con al centro una struttura simile ad anelli, il rachide.
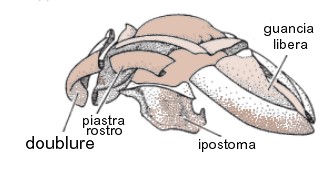
Il dorso era ricoperto da uno scudo protettivo che si rimboccava attorno ai margini del corpo con una piega chiamata doublure (immagine sopra); a livello del cephalon abbiamo quindi la doublure cefalica, mentre a livello delle pleure laterali abbiamo la doublure della pleura. Lo scudo o esoscheletro, era formato da una cuticola minerale bistratificata, calcitica a basso tenore di magnesio, suddivisa generalmente in uno strato esterno e in uno interno, che si differenziano per lo spessore e la matrice dei cristalli che li compongono.
Molti pori attraversavano, come dei canali, la cuticola dei trilobiti; probabilmente si trattava di peli sensoriali collegati al sistema nervoso centrale che trasmettevano informazioni dall'ambiente circostante. Presenti, sul dorso, anche tubercoli che probabilmente costituivano organelli sensoriali.
Il cephalon costituiva la porzione anteriore dell'animale e, come detto, deriva dalla fusione di più parti; la morfologia del cephalon è diversa da specie a specie e, grazie alle rare testimonianze fossili delle parti molli, sappiamo che i trilobiti avevano antenne sul capo e appendici ventrali (appendici cefaliche). La parte centrale del cephalon presentava una sorta di rigonfiamento, che si estendeva in parte al resto dello scudo toracico, noto con il termine di glabella, dalla forma variabile, ovvero da conica a cilindrica, e possedeva due guance laterali, le gene o genea. La glabella è delimitata lateralmente da due solchi assiali.
Le suture facciali dividono le gene in due parti distinte:
- fixigene (fixigenae) o guance fisse: la parte interna alle suture facciali, solidale alla glabella;
- ibrigene (librigenae) o guance libere: le aree esterne alle suture facciali, di forma semilunata, che sorreggono gli occhi.
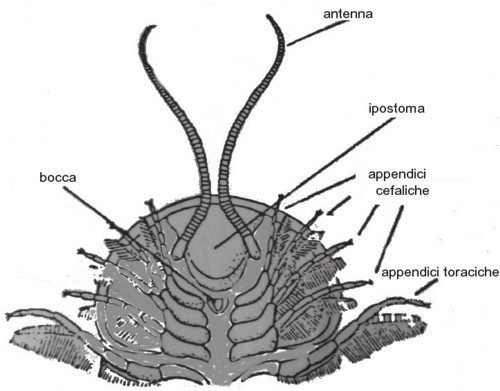
Il torace (thorax), era formato da un numero variabile di segmenti mobili (tergiti) e divisi ciascuno in tre parti, da due solchi longitudinali; la parte mediana di questi segmenti formava il rachide, mentre le pleure erano le zone laterali. La parte distale di queste ultime era estremamente variabile e dipendeva dalla specie; alcune terminavano con spine anche di dimensioni notevoli rispetto al copro dell'animale. I segmenti del torace erano fra loro articolabili così da permettere all'animale di arrotolarsi in caso di pericolo.
Il numero delle tergiti varia a due a 42; nella maggior parte delle specie descritte, il loro numero va da 8 a 15.
Il pigidio (pygidium), era costituito dalle tergiti non articolate e spesso fuse tra loro. Il numero di questi segmenti era variabile e talora il pigidio finiva posteriormente con una lunga punta caudale.
A seconda delle dimensioni relative del pygidium rispetto al cephalon, si distinguono:
- forme micropigie, in cui il pygidium è più piccolo del cephalon;
- forme isopigie, in cui il pygidium è comparabile con le dimensioni del cephalon;
- forme macropigie, in cui il pygidium è di dimensioni superiori rispetto al cephalon. Spesso il pygidium, in queste forme, era estremamente largo.
occasionalmente il pigidio terminava con una lunga spina detta spina assiale; nel sottordine Olenellina, eccezionalmente, il pigidio era formato da un'unica tergite, detta telson.
SVILUPPO
Questi organismi nella loro vita attraversavano vari stadi di sviluppo: nella prima fase, la larva (Protaspis) acquistava i metameri grazie a mute successive; in questa fase le larve erano piccolissime, da 0.3 a 1 mm circa. Potevano formarsi spine assiali, che poi erano perdute nei successivi stadi.
Nella seconda fase, la larva (Meraspis), presentava un corpo già abbastanza differenziato. Il cephalon era gia differenziato dal pigidio, ed era possibile osservare alcuni segmenti toracici. Tale fase è considerata terminata quando il numero dei segmenti è pari a quello dell'individuo adulto.
Nell'ultima fase (Holaspis), i trilobiti non subivano modificazioni morfologiche, ma la loro crescita continuava. Per cui esistono della stessa specie, fossili le cui dimensioni sono correlabili all'età raggiunta. Negli individui più grandi, il cephalon costituiva la maggior parte della superficie dell'esoscheletro.
Molti aspetti qui descritti in modo sommario, necessitano comunque di molti chiarimenti. La maggior parte dei fossili ci mostrano elementi disarticolati e dislocati su superfici
estese, depositati a caso dalle correnti e dalle attività di altri organismi. Esistono invece siti ove le parti fossili sono sovrapposte e mescolate, come il sito di Collingswood, Ontario (formazione di Collingswood).
Un esempio di ontogenesi, è riportato da Wikipedia.
L'aspetto relativo all'ontogenesi di questi artropodi è illustrata in un famoso contributo di Wittington, dal titolo Treatise of Invertebrate Paleontology, del 1959. Purtroppo non ci è stato concesso il permesso per riprodurre le bellisime sequenze fotografiche che mostrano l'ontogenesi di alcune specie di trilobiti. Pubblichiamo però le tavole di Barrande (1852) e di Bergeron (1899), esenti da copyright:
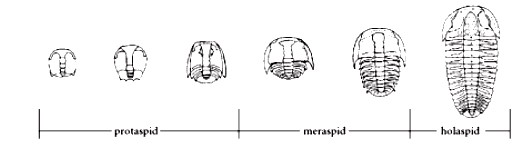
Sopra, Barrande (1852). Sao hiirsuta: stadio protaspis (x9); stadio meraspis (x7); holaspis (x0.5).
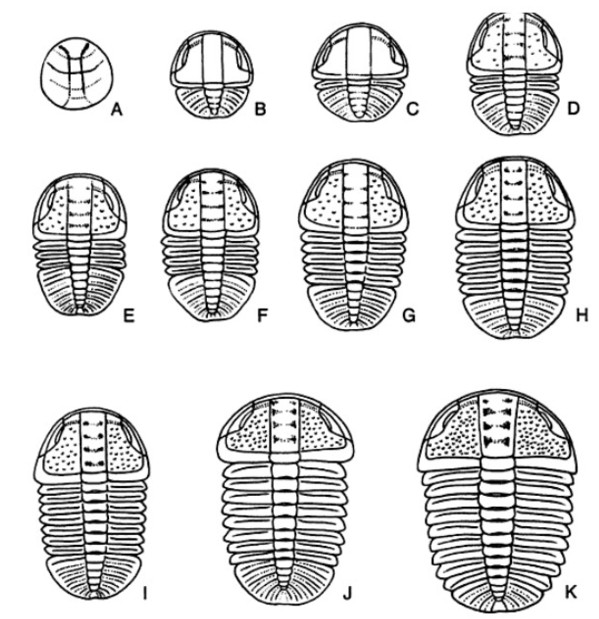
Sopra, Bergeron (1899). Ricostruzione ontogenesi Arthricoecephalus chauveaui. A protaspis (x26); B meraspis (x25); C meraspis (x23); D meraspis (x19); E meraspis (x17); F meraspis (x13); G meraspis (x15); H meraspis (x11); I meraspis (x9); J meraspis (x7); K holaspis (x7).
LA VISIONE: STRANE LENTI IN CALCITE

Occhio fossile composto di Erbenochile erbeni ottimamente preservato. Marocco, devoniano. L'immagine mostra un occhio schizocroico e nel dettaglio, in alto, si vede anche una sorta di struttura sporgente che fungeva da paraluce. Fonte immagine: Wikipedia.
Gli occhi di un qualsiasi animale possono dare preziosi suggerimenti sulla sua vita; purtroppo trattasi di tessuti molli che difficilmente giungono sino a noi attraverso i fossili. Ma gli occhi dei trilobiti erano diversi, si fossilizzarono molto bene perchè le lenti erano in calcite, ovvero dello stesso materiale con cui era costruito il loro esoscheletro.
I trilobiti avevano o occhi olocroici, con lenti esagonali fittamente stipate (sino a 15.000 per occhio), oppure occhi schizocroici, con lenti rotondeggianti e meno numerose, separate l'una dall'altra da materiale esoscheletrico.
La calcite ha una peculiarità: permette alla luce di attraversala, senza subire rifrazione, lungo uno degli assi cristallografici del minerale. Ogni lente dell'occhio dei trilobiti, permetteva alla luce di passare lungo un asse perpendicolare alla superficie della lente stessa e quindi, si è ricostruito il campo visivo dell'animale. I trilobiti osservavano al di sopra del fondale marino, ovvero lateralmente. Questo è vero sia per le specie con occhi olocroici che schizocroici, anche se le lenti di quest'ultimo tipo erano più sofisticate e permettevano la correzione dell'aberrazione sferica (Fortey R., 2004).
Alcune specie avevano adirittura occhi a torre (figura qui sopra)
con tanto di struttura sporgente per attenuare la luce proveniente dall'alto. Trilobiti più 'sofisticati' avevano poi occhi enormi e globulari; potevano ruotare le lenti in ogni direzione, quasi a 360° e pure verso l'alto. Altre specie di profondità persero invece gli occhi e divennero comopeltamente cieche. Sembra che forme cieche si siano evolute più volte durante la lunga storia evolutiva dei trilobiti.
Altre forme avevano occhi fissi e forse erano
pelagiche, ossia nuotavano liberamente senza aver vincoli con il fondale. Un esempio è Opipeuter (colui che ha lo sguardo fisso). Il loro esoscheletro dal peso ridotto era compatibile con la vita pelagica. L'ordovicano è stato il periodo in cui le forme pelagiche divennero dominanti e occupavano l'attuale nicchia ecologica del krill.
I trilobiti più 'strani' avevano enormi occhi ipertrofici e sporgenti, con lenti quadrate e non esagonali. Sono, non a caso, detti trilobiti ciclopigidi. Probbailmente vivevano in acque mesopelagiche, ovvero appena al di sotto della zona illuminata dal sole (150 - 200 metri). Altre specie erano epipelagiche evivevano dunque in prossimità della superficie degli oceani (es. Carolinites).
Dunque, anche se la maggior parte delle specie erano bentoniche, non sono rare forme liberamente natanti; alcune, a dispetto della loro scarsa idrodinamicità, avevano
carenature allungate, una sorta di rostro (es. Parabarrandia), che garantiva comunque loro una discreta velocità, sufficiente a predare piccoli crostacei primitivi.
Luoni Ottavio & Pierfederici Giovanni
SITOGRAFIA
ARTICOLI CORRELATI
- TRILOBITI - 2° parte
BIBLIOGRAFIA
- Whittington H.B. et al. - Arthropoda 1 Trilobita, Revised, Volume 1: Introduction, Order Agnostida, Order Redlichiida. in Treatise on Invertebrate Paleontology. The Geological Society of America & The University of Kansas Press, 1997. Originale 1959.
- Michael J. Benton,D. A. T. Harper - Introduction to paleobiology and the fossil record. Wiley.
- Richard A. Fortey - Trilobites and their relatives: contributions from the third international. Wiley.
- Richard A. Fortey - Vita da Trilobita. Le Scienze n. 436. 2004.
Questo articolo è protetto da Copyright © e non può essere riprodotto e diffuso tramite nessun mezzo elettronico o cartaceo senza esplicita autorizzazione scritta da parte dello staff di BiologiaMarina.eu.
Ideazione: Pierfederici Giovanni - Progetto: Pierfederici Giovanni, Castronuovo Motta Nicola, Guadagnino Marcello.
Prima Pubblicazione 31 Lug 2006 - Testi e immagini riproducibili secondo le specifiche Creative Commons. Le immagini dei Collaboratori detentori del Copyright © sono riproducibili solo dietro specifica autorizzazione.
Si dichiara, ai sensi della legge del 7 Marzo 2001 n. 62 che questo sito non rientra nella categoria di "Informazione periodica" in quanto viene aggiornato ad intervalli non regolari
XHTML 1.0 Transitional – CSS
