
BIOLOGIA MARINA SPECIALE
Stramenopile(i) E PSEUDOFUNGHI MARINI
Tags: funghi marini, pseudofunghi marini, traustochitridi, stramenopili, labirintulidi, oomiceti, sagenogene, sagenogentosoma, botrosoma, Quahog Parasite Unkonw, muffe mucillaginose, stomatocisti, statospore, ipnospora, ipnozigote, silicalemma, sinzoosporePer la prima volta, su Biologiamarina.eu scriviamo di funghi marini, o meglio, di pseudofunghi marini. In tal modo, senza pretesa di essere esaustivi, "proviamo" ad introdurre un argomento poco noto e non sempre oggetto di studio, anche in ambito universitario, soprattutto in Italia; online le informazioni in lingua italiana sono molto scarse, ma anche in inglese l’argomento si presenta lacunoso e frammentato, con l'esclusione di qualche cenno di sistematica e poco altro.
Innanzi tutto è indispensabile capire cosa si intende per "funghi marini" e perchè è stato coniato anche il termine composto "pseudofunghi".
La letteratura scientifica riporta diverse definizioni. La meno recente nella quale ci siamo imbattuti, durante la stesura di questo scritto, è quella di Johnson & Sparrow (1961), che definisce i funghi marini come "funghi capaci di crescere nell’acqua marina". La definizione è sicuramente semplicistica, per cui altri autori, negli anni, ne hanno proposte altre. Meyer (1968), definisce i funghi marini come "funghi capaci di crescere in acqua a determinate concentrazioni di cloruro di sodio", mentre Kohlmeyer & Jennings (1978), migliorano ulteriormente definendo "funghi marini tutti i funghi che crescono e sporulano obbligatoriamente in habitat marini o negli estuari".
Durante il 7° International Marine and Freshwater Mycology Symposium di Hong Kong, venne posta con una certa rilevanza la questione, solo apparentemente semplice, del tipo "Che cosa è un fungo marino?".
Ebbene, non si giunse ad una definizione univoca e accettata da tutti, ma furono proposte ed introdotti nuovi termini. Dunque i funghi marini sono "funghi alotolleranti, nativi di habitat esclusivamente marini e salmastri, capaci di crescere e sporulare in acque con un certo grado di salinità".
Non esiste una definizione più precisa e da tutti condivisa e, durante lo stesso Simposium, si delegò ai futuri ricercatori la "stesura della risposta definitiva", basata su conoscenze derivate dall'utilizzo di tecniche più moderne (allora agli albori e oggi di routine), come le analisi di biologia molecolare, ora maggiormente diffuse rispetto ad allora.
La definizione riportata da Wikipedia (http://en.wikipedia.org/wiki/Marine_fungi) (peraltro presa pari pari dal testo Marine Mycology, a Practical Approach, 2000) non sembra essere esaustiva e descrivere molto bene i fugnhi marini. Il testo più recente (Chandralata Raghukumar - Biology of Marine Fungi, 2012) non aggiunge molto alla questione e come altri, si limita a distinguere i funghi marini obbligati da quelli facoltativi.
I primi, secondo la definizione corrente, crescono e sporulano in mare e negli estuari, i secondi possono farlo anche in acque dolci e in habitat terrestri. Più in generale, le forme facoltative si accrescono e sporulano in ambienti terrestri e di acque dolci e solo secondariamente finiscono trasportate in mare. Attenzione, ad oggi non è stato dimostrato completamente che i funghi marini classificati come "obbligati" completino l’intero ciclo vitale in acque salate (Darry, 2009).
Dunque, molto importante, i funghi marini non sono da intendersi come un gruppo tassonomico, piuttosto si tratta di un gruppo di organismi eucarioti "fisiologicamente ed ecologicamente definito".
Sono note 444 specie di veri funghi marini: 360 ascomiceti, 10 basidiomiceti, 74 funghi mitosporici. Jones et al. riportano invece, nel loro lavoro più recente, circa 530 specie.
FUNGHI E PSEUDOFUNGHI
Vediamo adesso di esaminare la questione "veri funghi" e "pseudofunghi"; si tratta peraltro di introdurre un breve escursus storico, tornando indietro nel tempo, quando Haeckel, discepolo di Darwin, creò il regno dei Protisti che raggruppava tutti gli organismi microscopici, quindi batteri, alghe, protozoi e microfunghi.
Negli anni '50 del '900, l'avvento del microscopio elettronico permise di scoprire nuove particolarità, ovvero che i batteri sono procarioti con cellule prive di nucleo e dalla struttura cellulare, in generale, molto diversa da quella delle cellule eucariote.
Nel 1969 la Whittaker
assegna i funghi ad un regno separato, pur sapendo che il gruppo era polifiletico, poiché non era identificabile un unico progenitore comune.
Dunque siamo arrivati al punto in cui gli organismi viventi sono sudivisi in due domini, i procarioti e gli eucarioti, almeno sino a quando apparve evidente che certi batteri presentavano caratteristiche talmente diversi da dover creare un terzo dominio, quello degli Archea. Nel tempo, il complesso albero filogenetico si complicò e, nel caso dei funghi, si giunse ad una suddivisione che è rimasta valida, per ragioni sia storiche che pratiche, sino a pochi anni fa.
Dunque il gruppo dei veri funghi, o miceti, è caratterizzati da attributi cellulari e biochimici tipici (crescita ifale, nutrizione per assorbimento, parete cellulare chitinosa, nuclei aploidi, microtuboli sensibili a benzimidazoli e griseofulvina ecc..). A tale gruppo appartengono molti microrganismi marini microscopici (chitridi o chitridiomiceti, zigomiceti, basidiomiceti, ascomiceti e deuteromiceti);
segue il gruppo delle muffe mucillaginose, che presenta stadi di sviluppo protoplasmatici, con cellule prive di parete, dunque manca la chitina sostituita dalla galattosamina. La somiglianza con i funghi è data solo dal fatto che producono spore;
segue il gruppo degli oomiceti, diversi dai veri funghi in quanto con nuclei diploidi e parete cellulosica, nonché diversi per altre caratteristiche che non esaminiamo in questa sede.
Nel tempo furono coniati diversi termini, per esempio le muffe mucillaginose divennero "funghi affini ai protozoi", mentre gli oomiceti mutarono in "funghi affini alle alghe eteroconti". In sintesi, molti eucarioti microscopici rimasero oggetto di studio dei micologi, mentre altri erano studiati dai ficologi (studio delle alghe), poichè considerati, appunto, delle alghe. Ecco perchè, ancora oggi, alcuni organismi che funghi non sono, continuano ad essere considerati tali.
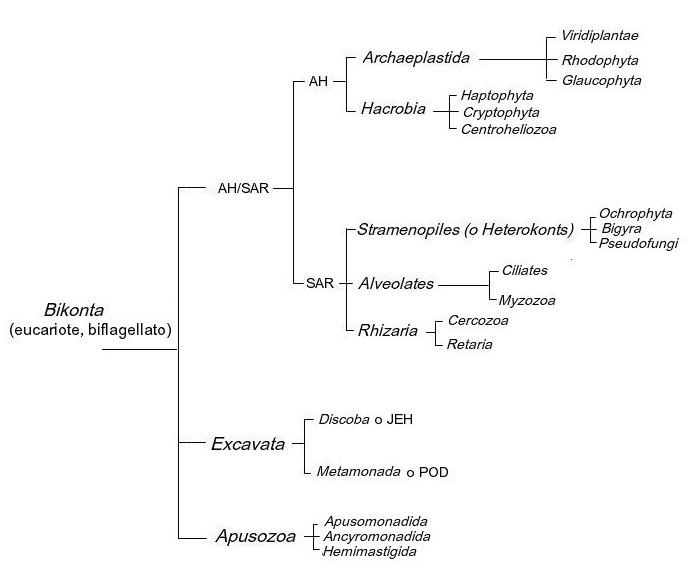
Passa altro tempo e gli eucarioti vengono divisi in sei supergruppi:
- Amebozoa;
- Opisthokonta;
- Archaeplastida, ovvero il vecchio Regno Plantae; comprende alghe Glaucocystophyta, Rhodophyta e Chloroplastida;
- Chromalveolata, organismi eterotrofi che avrebbero acquisito i cloroplasti da un endosimbionte autotrofo, forse un'alga rossa. Alcuni poi, hanno o avrebbero perso i cloroplasti divenendo di nuovo eterotrofi (eterotrofia secondaria), altri avrebbero invece riacquisito i cloroplasti persi in precedenza (endosimbiosi terziaria). Alcuni di questi protozoi non digeriscono le alghe microscopiche di cui si nutrono, ma di esse conservano i cloroplasti, spesso avvolgendoli in membrane pluristratificate. Tra questi ricordiamo: Cryptophyta, Haptophyta, Alveolata e Stramenopili detti anche Heterokonta;
- Excavata, protozoi caratterizzati da una escavazione detta citostòma o citofaringe. Sono eterotrofi primari (osmotrofici, saprofiti, fagotrofi) perchè non hanno mai acquisito cloroplasti, oppure eterotrofi secondari perchè gli hanno persi successivamente. Le poche specie note con cloroplasti sono fototrofe e sembrano essere endosimbionti secondari (hanno perso e poi riacquisito i cloroplasti). A questo taxon appartengono le Euglenophyceae.
- Rhizaria, organismi ameboidi con pseudopodi. Comprendono i taxa Chlorarachiniophyceae e Cryptopphyceae.
Oggi, grazie a studi di filogenesi molecolare,
anche se ancora lontani da una classificazione definitiva, questo supergruppo con sei taxa è stato ulteriormente smembrato. Nell'immagine sottostante vediamo che Rhizaria, Alveolata e Stramenopila sono finiti accumunati in un ramo dell'albero filogenetico diverso da quello delle Cryptophyta e Haptophyta.
Per non appensatire il discorso, da adesso in poi la classificazione utilizzata è quella riportata nello schema sottostante.
Ci occuperemo, nello specifico, delle Stramenopile, vasto gruppo di organismi, quasi tutti eterotrofi, un tempo studiati in parte dai micologi e in parte dai ficologi perchè rispettivamente, considerati in parte funghi e in parte alghe. Oggi sappiamo che non è così.
Stramenopiles, LABIRINTULOMICETI E TRAUSTOCHITRIDI
Gli eteroconti o stramenopili (dal latino stramen = filo, fuscello di paglia e pilous = capello, pelo), costituiscono un cospicuo gruppo di organismi eterogenei, che sono comunque raggruppati sulla base non solo di affinità morfologiche, ma soprattutto molecolari. Le stramenopile (o gli stramenopili, in letteratura abbiamo trovato entrambe le accezioni maschili e femminili), possiedono tutte, almeno in un momento del loro ciclo vitale, due flagelli.
I flagelli sono eteroconti (cioè sono diversi), ed entrambi inseriti al polo anteriore della cellula. Il più lungo dei flagelli è pleuronematico ed è orientato anteriormente, l'altro, il più corto, è anematico ed orientato posteriormente, inoltre è dotato di rigonfiamento basale ove è situata una macchia oculare.
Il flagello pleuronematico, possiede su ambedue i lati, piccoli peli rigidi tubulari tripartitici, chiamati mastigonemi (indicati spesso con l'acronimo TTHs = Tubular Tripartite Hairs); delle tre parti dei mastigonemi, si distingue la porzione basale, un manico tubulare ricoperto da peli molto più piccoli e, infine, uno o più peli terminali; il secondo flagello, anematico, è completamente spoglio. Da qui il termine eteroconte, che significa appunto, flagelli diversi. Alcune stramenopile hanno perso i mastigonemi nel corso della loro evoluzione. Altra caratteristica delle stramenopile sono la mitosi aperta e la presenza di mitocondri con creste tubulari.
La classificazione delle stramenopile è tutt'altro che definitiva. Thomas Cavalier-Smith racchiude il taxa nell'infraregno Heterokontophyta, che comprende diversi phyla e diverse classi; noi invece riportiamo la classificazione di Ingvild Riisberg et al., basata su due phyla eterotrofi e uno autotrofo. Entrambe le classificazioni comunque sono ad oggi accettate. Riisberg et al. non riportano alcune classi che invece riporta Cavalier-Smith (come Synurophyceae, Chrysomerophyceae ecc..).
- phylum Ochrophyta [sin. Chromophyta], che comprende il subphylum Phaeista e Khakista (alghe bruno-dorate note come crisoficee, le eustigmatoficee, le rapidoficee ecc...). Organismi autotrofi.
- phylum Bigyra (Bicosoecida, Blastocystis e Labyrinthulida). Organismi eterotrofi.
- phylum Pseudofungi (o Heterokontimycotina). Organismi eterotrofi.
Entrambi i phyla eterotrofi annoverano specie che non possiedono il gene rblc (che codifica per una delle subunità della rubisco).
Il phylum Pseudofungi è interessante e comprende microrganismi largamente diffusi in mare, oltre i gia citati oomiceti (= fungo a uovo). Anche per loro non vale più l’appartenenza al gruppo dei veri funghi, anche perchè, come scritto sopra, hanno cellule con pareti in cellulosa e beta-glugano, oltre ad essere dipliodi per l’intero ciclo vitale. Durante la riproduzione sessuale producono una cellula uovo molto grande, detta oogonio, che può essere fecondata o da una cellula spermatica o da una cellula gametica più piccola detta anteridio, dopo il quale si forma uno zigote.
Al gruppo degli oomiceti appartengono i peronosporomiceti che formano ammassi cotonosi su alghe e animali morti. Un famoso porosporomicete è Phytophthora infestans, che nel 1845 causò la carestia d’Irlanda, poiché distrusse l'intera produzione di patate. Torneremo in un altro momento a scrivere sugli oomiceti.
Alle stramenopile appartengono anche i labirintulomiceti o labirintulidi. Si tratta di organismi unicellulari un tempo classificati come veri funghi, sia per affinità morfologiche che per ovvie ragioni storiche gia avallate sopra, ma la scoperta di zoospore con due flagelli ha portato alla revisione della loro sistematica.
Lo stadio più comune del loro ciclo vitale, non flagellato, consta di cellule fusiformi che formano colonie complesse entro una rete ectoplasmatica, all’interno della quale si spostano con movimenti ameboidi. Tale rete, fortemente anastomizzata, è formata da materiale di origine ectoplasmatico, ricco di fibre actino-simili e calcio-dipendenti.
L’essudato ectoplasmatico è privo di organelli ectoplasmatici perchè filtrato da strutture caratteristiche, note con i nomi di botrosoma o sagenogene (sagenogentosoma). Tali strutture, dense agli elettroni, sono dislocate sulla parete cellulare e appaiono leggermente invaginate, di fatto separano la matrice ectoplasmatica dalle componenti interne della cellula.
Le cellule si muovono entro la rete ectoplasmatica con movimenti ameboidi e si riproducono per scissione binaria o per mezzo di zoospore biflagellate eteroconti. I labirintulidi sono osmotrofici e producono enzimi digestivi extracellulari, che li avvicina appunto, almeno fisiologicamente, ai veri funghi.
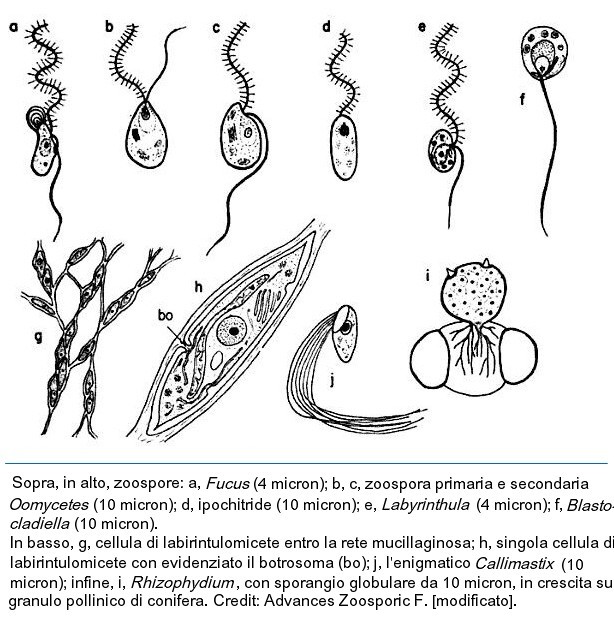
Sono facilmente isolabili da alghe morte e detriti galleggianti. Il loro ciclo vitale può essere sintetizzato come segue: zoospore o cisti generano cellule elissoidi, le quali producono la rete ectoplasmatica prima descritta. Queste cellule, dette cellule trofiche, si aggregano tra loro a formare colonie e, successivamente, uno sporangio dal quale si formano sporociti che producono zoospore.
Gli sporangi possono aggregarsi e formare sori. Singole cellule dello sporangio posso formare cisti e dar vita a nuove cellule trofiche.
Le colonie sono formate da un centinaio di cellule, di dimensioni da 8-25 x 3-8 micron. Man mano che si accresce la colonia, aumentano le divisioni mitotiche e le cellule centrali tendono a spostarsi verso la periferia, muovenmdosi di alcuni micron ogni minuto.
I labirintulomiceti sono ubiquitari nei mari e negli estuari. Si trovano associati a detriti galleggianti e ai sedimenti marini; alcune specie sono state isolate da ferite di animali, soprattutto nei cefalopodi e nei nudibranchi.
Alcune specie sono responsabili anche di patologie che colpiscono i molluschi. Per esempio Labyrinthuloides haliotidis colpisce alcune specie di abalone (Haliotis kamtschatkana e H. rufescens); un'altra specie senza nome, responsabile della malattia QPX (Quahog Parasite Unkonw) è stata isolata in Mercenaria mercenaria e in M. mercenaria var. notata, molluschi molto diffusi in nord America chiamati volgarmente Quahog. Analisi molecolari, hanno sino ad ora permesso di stabilire che l'agente eziologico della malattia è forse un protista traustochitride. Schizochytrium sp e Ulkenia ammoeboidea, sono state isolate da lesioni ulcerative precedute da alterazione dei cromatofori, in molluschi cefalopodi, in particolare polpi. Da alcuni calamari (Illex illecebrosus), con evidenti lesioni ai tessuti branchiali, sono state isolte diverse specie di traustochitridi.
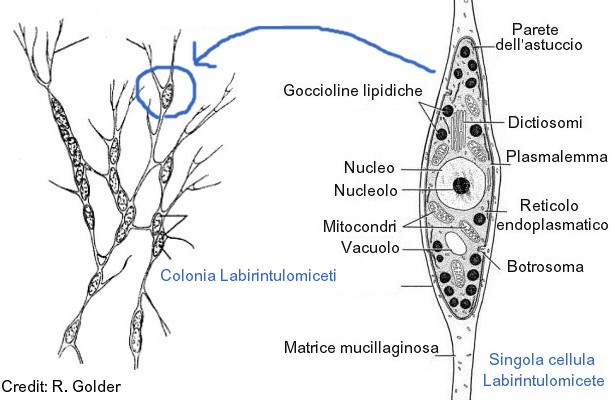
Sopra, a sinistra colonia di Labirintulomiceti e, a destra, rappresentazione di una cellula tipica.
Altre stramenopile sono state a lungo studiate dai ficologi e dai micologi. Si tratta perlopiù di protozoi eterotrofi e, in alcuni casi, autotrofi. Nel paragrafo successivo riportiamo alcune brevi caratteristiche di ogni taxa elencato.
CLASSIFICAZIONE STRAMENOPILE
La sistematica delle stramenopile, come detto, è in continua revisione. Quella che presentiamo è dedotta dal lavoro di Ingvild Riisberg et al., Seven Gene Phylogeny of Heterokonts, Protist, Vol.160, 191-204, maggio 2009.
Phylum Ochrophyta [sin. Chromophyta]. Tale phylum, secondo diversi lavori, tra cui quelli di Cavalier-Smith, comprendeva diverse classi, alcune delle quali confermate dal lavoro di Riisberg et al.; altre invece sono state escluse, altre ancora non sono state confermate ma comunque incluse perchè moderatamente supportate dalle analisi di filogenesi molecolari.
Questo phylum, secondo Riisberg et al., comprende:
subphylum Khakista
- Classe: Pelagophyceae: alghe prevalentemente marine e pelagiche, possono essere flagellate o prive di flagelli, formare colonie filamentose e a volte proliferare in massa, originando le cosiddette brown tides.
- Classe: Dictyochophyceae (ex Silicoflagellate): alghe flagellate e talvolta ameboidi; alcuni generi presentano cellule nude, altri hanno cellule ricoperte da scaglie e, infine, il genere Dictyocha, presenta cellule con scheletro siliceo. Queste ultime sono note sin dal Cretaceo, e si trovano numerose in sedimenti fossili ben conservati. Le D. vivono sia in acque dolci che marine.
- Classe: Bacillariophyceae: a tale classe appartengono le diatomee, diffuse sia in acque dolci che salate di tutto il globo. Tale gruppo è complesso e in parte è stato discusso qui, per gli altri aspetti invece, torneremo con altri contributi.
subphylum Phaeista
- Classe: Phaeophyceae: note come alghe brune, sono prevalentemente marine e possono raggiungere dimensioni ragguardevoli; per esempio il kelp che ricopre estese superficie, raggiunge dimensioni di svariati metri. Sono tutte pluricellulari e accumulano al loro interno grandi quantità di iodio e bromo. I gameti e le zoospore presentano i tipici flagelli delle stramenopile. I flagelli terminano entrambi con un acronema (parte sottile con solo due microtubuli). Alghe complesse, saranno trattate in modo più dettagliato in uno dei prossimi articoli.
- Classe: Xanthophyceae: note come alghe giallo-verdi, presentano morfologia coccale, sifonale, ameboide o flagellata; parete cellulosica impregnata a volte di silice o carbonato di calcio. Quasi tutte di acque dolci, formano feltri sulle rive di laghi e fiumi (sono epifite, ovvero aderiscono a substarti di vario tipo). Si riproducono asessualmente e formano zoospore e, in condizioni sfavorevoli, ipnospore endogene, racchiuse da una parete silicea a due valve. Rara la riproduzione sessuale, con isogamia e formazione di ipnozigoti. Nelle forme sifonate come Vaucheria, si formano, all'interno di sporocisti, spore multinucleate e multiflagellate dette sinzoospore o zoospora composta (forse derivata dall'unione di più zoospore) che, una volta libere, nuotano per fissarsi poco dopo al fondo originando forme cenocitiche. Vaucheria si riproduce anche sessualmente originando all'interno del citoplasma, gametocisti maschili e femminili isolate da setti appositi, i quali contengono rispettivamente spermatozoidi biflagellati e una sola oosfera uninucleata. Dopo la fecondazione si forma una spessa parete e lo zigote diviene quiesciente.
- Classe: Phaeotamnophyceae: alghe di acque dolci racchiuse in tale classe nel 1998 da Bailey, sulla base di studi molecolari e biochimici. Sono un sister gruop delle Xanthophyceae.
- Classe: Crysophyceae: note come alghe dorate per il color giallo oro del cloroplasto, sono alghe microscopiche unicellulari eterogenee; la morfologia è variabile, possono essere nude, ameboidi, ricoperte di scaglie silicee, avere parete cellulosica o alloggiare in loriche polisaccaridiche dalla forma particolare (vaso, calice ecc..), possono essere impregnate di ferro o manganese. Possono formare spore endogene dette stomatocisti o statospore, variamente ornate con protusioni più o meno elaborate, con parete silicizzata come quella delle diatomee, derivante dal silicalemma che si forma nel citoplasma periferico; presentano, le cisti, un'apertura occlusa da una sorta di tappo a forma di cono. Tale cisti sono a volte associate ad ipnozigoti. Sono prevalentemente di acque dolci, soprattutto di acque fredde e oligotrofiche, ma sono rappresentate anche da alcuni generi marini. Si riproducono asesualmente (rari i casi di riproduzione sessuale); ciclo monogetetico aploide.
- Classe: Eustigmatophyceae: alghe diffuse sia in acque dolci che salate, si caratterizzano per la presenza di una macchia oculare o stigma esterna al cloroplasto. Quest'ultimo ha un pirenoide peduncolato non espresso nelle zoospore.
- Classe: Pinguiophyceae: microalghe flagellate o non flagellate note per il loro peculiare contenuto di omega-3 (latino pingue = grasso). Sono un sister gruop delle Xanthophyceae.
Phylum Bigyra. Questo phylum, come quello pseudofungi che vedremo, era stato escluso dal taxa Ochrophyta come dimostrano alcuni lavori recenti, per esempio Cavalier-Smith & Chao (2006) e Leipe et al. (1994, 1996). Le analisi di filogenesi molecolari collocano comunque molto vicini tra loro Bigyra, Pseudofunghi e Ochrophyta. Il phylum Bigyra comprende oltre 80 specie e spesso è così suddiviso:
- Classe: Blastocystea (ex subphylum Opalinata): protozoi parassiti del tratto gastrointestinale di molte specie animali, uomo compreso. La specie Blastocystis hominis è stata identificata come responsabile della blastocistosi.
- Classe: Labyrinthulea ( ex subphylum Sagenista). Comprende Labyrinthulidae e Traustochytridae.
- Classe: Bicosoeciea sin. Bicoecea: piccolo gruppo di protozoi non parassiti e privi di coloroplasti, sul quale esistono poche informazioni. Comprende i generi Bicosoeca, Cafeteria, Pseudobodo, Siluania e Symbiomonas.
Phylum Pseudofungi (o Heterokontimycotina). Phylum che comprende Oomycota e Hypochytriomycetes, nonchè la specie Developayella elegans.
- Classe: Oomycetes
- Classe: Hyphochytridiomycetes
Per il momento l'introduzione alle Stramenopile termina qui. Se avete dubbi, domande, se notate imprecisioni o desiderate integrare l'argomento con ulteriori informazioni, non esitate a contattarci; l'argomento è complesso e troppo poco dibattuto, quindi ben venga qualsiasi ulteriore notizia e informazione.
AGGIUNGI UN COMMENTO [Se non desideri visualizzare il tuo commento sul tuo profilo Facebook, ricordati di togliere la spunta della ceckbox]
ARTICOLI CORRELATI
BIBLIOGRAFIA
- Cavalier-Smith T., Chao E.E.Y. - Phylogenyandmega- systematics ofphagotrophicheterokonts(kingdomChro- mista). J.Mol.Evol. 62: 388-420. 2006.
- Darryl L. Felder, David K. Camp - Gulf of Mexico Origin, Waters, and Biota:Volume I. Texas A&M University Press. 2009.
- Ingvild Riisberg et al. Seven Gene Phylogeny of Heterokonts. Protist, Vol. 160, 191-204. 2009.
- Jones, E.B.G., J. Sakayaroj, S. Suetrong, S. Somrithipol and K.L. Pang. 2009. Classification of marine Ascomycota, anamorphic taxa and Basidiomycota. Fungal Divers. 35: 1–187.
- Leipe D.D., Wainright P.O., Gunderson J.H., Porter D., Patterson D.J., Valois F., Himmerich S., Sogin M.L. - The Stramenopiles from a molecula rperspective-16S-like ribosomal-RNA sequences from Labyrinthuloides minuta and Cafeteria roenbergensis. Phycologia 33: 369—377. 1994.
- Leipe D.D., Tong S.M., Goggin C.L., Slemenda S.B., Pieniazek N.J., Sogin M.L. - 16S-liker DNA sequences from Developayella elegans, Labyrinthuloides haliotidis, and Proteromonas lacertae confirm that the stramenopiles area primarily heterotrophic group. Eur. J. Protistol 32: 449-458. 1996.
Questo articolo è protetto da Copyright © e non può essere riprodotto e diffuso tramite nessun mezzo elettronico o cartaceo senza esplicita autorizzazione scritta da parte dello staff di BiologiaMarina.eu.
Ideazione: Pierfederici Giovanni - Progetto: Pierfederici Giovanni, Castronuovo Motta Nicola.
Prima Pubblicazione 31 Lug 2006 - Le immagini dei Collaboratori detentori del Copyright © sono riproducibili solo dietro specifica autorizzazione.
Si dichiara, ai sensi della legge del 7 Marzo 2001 n. 62 che questo sito non rientra nella categoria di "Informazione periodica" in quanto viene aggiornato ad intervalli non regolari
XHTML 1.0 Transitional – CSS
