
I PORIFERI Parte I

Aplysina sp. Immagine di M. Guadagnino
I Poriferi (dal latino porus = poro e ferre = portare), sono organismi appartenenti al sottoregno dei Parazoi e sono tutti sessili. Li troviamo sia in mare che nelle acque dolci, e sono carattrizzati dall'avere un innumerevole varietà di forme, che li rende facilmente confondibili con altri organismi bentonici. Sono definiti organismi sopspensivori, anche se alcune spugne possono spostarsi attraverso complesse migrazioni cellulari. Sono state classificate sino ad oggi oltre 9.000 specie di spugne.
Il livello organizzativo dei Poriferi è abbastanza semplice, ma al tempo stesso unico, il che rende difficoltoso risalire ai rapporti filogenetici con altri phyla animali. A differenza di tutti gli altri organismi multicellulari che hanno due tipi di tessuti, quello epiteliale e quello parenchimatico, nelle spugne si trova solo un tessuto strutturalmente e funzionalmente semplice. Anche la matrice extracellulare (ECM) è scarsa. 
Sono privi di una simmetria particolare, e in natura sono presenti strutture sacciformi, a coppa, arboescenti, incrostanti ecc... Per semplicità descriveremo ora la struttura più semplice, detta ascon (dal greco askós, sacco), o di tipo asconoide, che si presenta come un sacco verticale, con una cavità interna detta sacco ventrale, a pareti sottili.
Cominciamo dalle pareti del sacco. Sono costituite da due strati cellulari delimitanti una massa gelatinosa detta mesoglea o mesoilo. Lo strato esterno è detto strato dermale o pinacoderma, formato da cellule dette pinacociti che si presentano sottili e appiattiti. Lo strato interno è detto strato gastrale o gastroderma o anche coanoderma, ed è costituito da cellule dette coanociti lassamente connessi tra loro.
La camera centrale è detta atrio, ma è nota anche come spongocele o coanocele, che riceve acqua attraverso dei minutissimi pori detti ostii, localizzati sulla superficie della spugna che costituisco quindi i pori inalanti. Questi ultimi derivano da cellule specializzate del pinacoderma dette porociti. Sviluppandosi, tali cellule si allungano e si arrotolano come un tubo, attraversando l'intero spessore della spugna sino a originare appunto un'apertura. Se l'apertura è formata da un solo porocito "arrotolato" si parla appunto di ostio, se è formata da più porociti prende il nome di poro dermale.
L'acqua all' interno dello spongocele viene mossa grazie ai flagelli dei coanociti,
e viene espulsa attraverso un'apertura esalante detta osculo, che nelle spugne tipo ascon si trova all'apice della spugna stessa.
Tra i due strati come detto abbiamo il mesoilo, ricco di una matrice acellulare gelatinosa e colloidale, dove sono presenti cellule dette archeoblasti, capaci di differenziarsi in altri tipi cellulari sulla base delle necessità. Possono differenziarsi in amebociti, che distribuiscono le sostanze nutrienti, e in gametociti, che descriveremo più avanti. Alcune cellule producono le spicole, che sono delle strutture di sostegno localizzate sempre nella mesoglea. Sono utilizzate per classificare le varie specie di spugne.
I limiti di una struttura asconoide sono le dimensioni. Infatti il volume dell'acqua all'interno della mesoglea cresce secondo un rapporto cubico rispetto all'altezza del corpo,
e la superficie interna cresce secondo un rapporto quadratico, sempre rispetto all'altezza, per cui non è possibile per strutture così semplici raggiungere dimensioni elevate. Difatti molte spugne si presentano ocme asconoidi nel primo stadio di crescita (olinto), poi cambiano forma.

Spirastrella cunctatrix. Immagine di M. Guadagnino
Allora non resta che ripiegare la superficie corporea senza notevoli incrementi di mole. Si ha a che fare in questo caso con strutture dette siconoidi, o spugne di tipo sycon (dal greco sykon, spugna). Il ripiegamento della superficie corporea verso l'interno porta alla realizzazione dei canali inalanti, tapezzati internamente dai pinacociti e comunicanti con l'esterno attraverso gli ostioli o pori dermali, a seconda se circondati da un'unica o più cellule. I pinacociti che rivesto i canali interni prendono il nome di endopinacociti. Dall'evaginazione dello strato dermale si originano invece i canali esalanti o canali radiali, tapezzati dai coanociti flagellati. Nelle spugne sycon quindi, i coanociti non sono più presenti nello spongocele, peraltro molto ridotto, bensì solamente entro i canali radiali.
Il vantaggio delle spugne siconoidi è quello di aumentare notevolmente la superficie di scambio con l'acqua, e i canali radiali sono lunghi tanto quanto lo spessore del corpo.
I canali radiali comunicano con lo spongocele attraverso aperture dette apopili,
mentre i canali inalanti favoriscono l'ingresso di acqua attraverso i prosopili. Tali aperture sono degli spazi che si aprono tra cellule contigue (spazi intracellulari).
Quindi il percorso dell'acqua è questo: poro inalante -> canale inalante -> prosopilo -> canale radiale o esalante o flagellato -> apopilo -> spongocele -> osculo.
La figura sottostante permette di capire come è fatta la struttura delle spugne ascon, sycon e leucon:
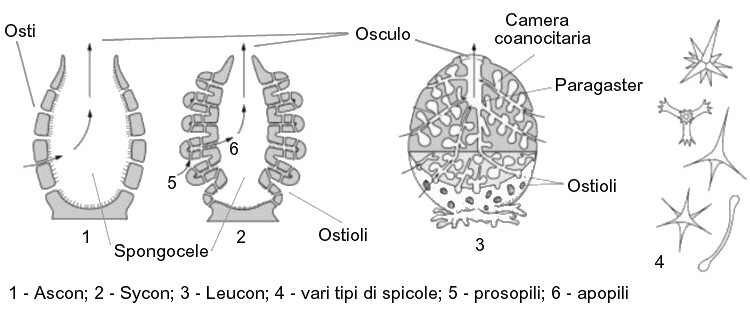
Il coanoderma può mantenersi liscio (condizione asconoide), ripiegarsi (condizione siconoide) e infine può suddividersi in numerose camere flagellate, quest'ultima è la condizione leuconoide, e le spugne sono dette di tipo leucon (dal greco leukós, bianco). Si tratta nel grado di complessità più alto raggiungibile dalle spugne. Dalla ulteriore estroflessione dei canali radiali prendono origine le camere coanocitarie, che permettono alla spugna di incrementare ulteriormente la superficie di scambio con l'acqua. Lo spongocele si riduce ad un canale centrale, il paragaster. Le camere coanocitarie sono solitamente sferiche e tra esse e il canale radiale si trovano gruppi di cellule dette cellule centrali, che regolano il flusso dell'acqua. Anche il mesoilo si differenzia in una regione corticale esterna (cortex) contenente gli elementi dello scheletro e priva di camere, e in una interna detta coanosoma che ospita le camere coanocitarie.
Non tutte le spugne passano attraverso i tre stadi descritti, alcune infatti mantengono la condizione asconoide e siconoide anche da adulte, come le spugne Calcarea, altre ancora sono completamente diverse da quelle ora descritte.

Colonia di Haliclona mediterranea, Immagine di M. Guadagnino
LE CELLULE DELLE SPUGNE
Solo alcuni tipi cellulari delle spugne sono stati caratterizzati sia funzionalmente che morfologicamente, della maggior parte infatti non si conoscono le funzioni fisiologiche. Qui riportiamo un riassunto dei tipi descritti sino ad oggi:
Pinacociti: Le spugne non formano veri tessuti epiteliali in quanto i pinacociti sono privi di desmosomi. I pinacociti si presentano in genere appiattiti e ricchi di granuli. A volte accanto a pinacociti appiattiti si alternano pinacociti a forma di T, è il caso del pinacoderma delle spugne calcaree. Come detto si hanno i pinacociti esterni, o esopinacociti, e quelli dei canali interni o endopinacociti che formano l'endopinacoderma. Quelli interni sono meno addossati gli uni agli altri, e svolgono anche una funzione fagocitaria. Infine abbiamo anche i basopinacociti. Vediamone le differenze:
- gli esopinacociti nella maggioranza delle spugne si presentano a forma di T, e in alcuni casi producono delle estroflessioni fusiformi che si connettono lassamente con le cellule contigue. In alcune specie come Hippospongia communis, Ephydatia fluviatilis, Sycon coactum, Oscarella lobulairs e O. tubercolata le zone di contatto degli esopinacociti presentano zone dense agli elettroni. Nelle Homoscleromorpha gli esopinacociti sono associati a dense fibrille e collagene di tipo IV, oltre alla tenascina. Caso particolare, sempre in Homoscleromorpha gli esopinacociti sono flagellati e possono produrre spicole. Il nucleo in genere si presenta ovale e in posizione centrale, mentre la cromatina è uniformemente distribuita. Sono ben evidenti i mitocondri e i lisosomi. Contengono anche fagosomi e vacuoli positivi ai trigliceridi. Il nucleolo a volte è presente a volte non risulta visibile. Come detto sono strettamente addossati tra loro, e mancano vere giunzioni cellulari, tuttavia le porzioni delle pareti cellulari a contatto tra loro sono specializzate dal punto di vista ultrastrutturale.
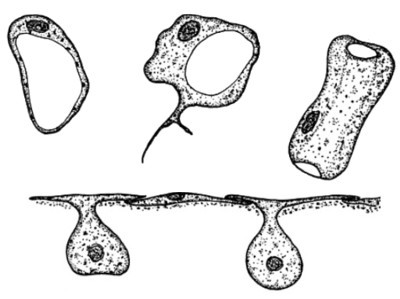
Sopra, in alto due porociti in sezione, a destra un porocito avvolto a formare un ostio. Sotto, esopinacociti, appiattiti e addossati gli uni agli altri e formanti una struttura detta a T. E' visibile il nucleo. Fonte: Patricia R. Bergquist - Sponges. 1979
- gli endopinacociti si distinguono dagli esopinacociti per la loro localizzazione, rivestono infatti i canali interni, e si presentano morfologicamente subsferici o fusiformi. Si suddividono in prosopinacociti che rivestono i canali inalanti e in apopinacociti che rivestono quelli esalanti. A volte si estendono sino al mesoilo. Inoltre non sono strettamente addossati tra loro e solitamente distano 10-30 nanometri. Il nucleo si presenta ovale e la cromatina dispersa. Raramente presentano delle cilia, soprattutto se dislocati nelle canali più grandi (per esempio in Hippospongia e Oscarella). Infine la loro area contrattile è ridotta rispetto agli esopinacociti. . In tutte le Homoscleromorpha e in alcune Demospongiae gli endopinacociti sono flagellati, e nel caso specifico della spugna Tethya lyncurium la presenza dei flagelli è associata alla movimentazione dell’acqua. Sempre nelle spugne Homoscleromorpha e alcune Demospongiae sono ricoperti da uno strato glicoproteico (glicocalice). Nella spugna Oscarella gli endopinacociti sono uniti in una zona di giunzione che prende il nome di zonula adhaerens (Ereskovsky, 2009).
- i basopinacociti formano l'epitelio basale o squamoso e si presentano appiattiti. Producono fibrille di collagene e polisaccaridi di varia natura, che costituiscono nel complesso la lamina basale della spugna, indispensabile per l'ancoraggio al substrato. Nelle specie di acqua dolce possono produrre filopodi che estendono nell'ambiente esterno circostante. Svolgono quindi un ruolo trofico e un ruolo osmoregolatorio. Il nucleo si presenta ovale, la cromatina dispersa e il nucleolo è ben visibile, stranamente, nelle sole specie di acqua dolce. Anche i fagosomi sono presenti nelle specie di acqua dolce. Presentano numerosi mitocondi e vacuoli contrattili che eliminano l'acqua in eccesso. Nella demospongia coralligena Acanthochaetetes wellsi i basopinacociti producono un massivo scheletro calcareo, mentre nella specie Ephydatia muelleri sono state descritte delle strutture simili ai desmosomi (Pavans de Ceccatty, 1986).
I pinacociti mancano nelle spugne Esattinellidi.
Porociti: si presentano (vedere figura sopra) fusiformi e cilindrici, tipo un manicotto e derivano dagli esopinacociti. Il loro nucleo si presenta ovale, la cromatina aggregata in patch, il nucleolo è assente. In alcune specie di spugne di acque dolci i vacuoli sono contrattili. Producono fillopodi. Formano gli ostii nelle spugne asconoidi e possono aprire, chiudere e modulare l'apertura a seconda delle necessità. I porociti in Haplosclerida e Demospongiae non sono a manicotto bensì appiattiti e presentano un’apertura centrale o periferica, e funzionano come uno sfintere.
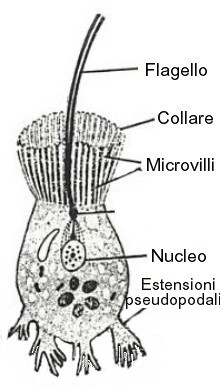 Coanociti: sono cellule flagellate fondamentali per la nutrizione delle spugne. Nelle spugne asconoidi tappezzano lo spongocele, mentre nelle siconoidi solamente i canali radiali. Formano quello che viene detto coanoderma. Il flagello è circondato da un collaretto citoplasmatico non compatto, bensì formato da un insieme di strutture più piccole addossate tra loro, che nella letteratura anglosassone sono detti microvilli o tentacoli citoplasmatici. Sono presenti 20-55 microvilli e il loro numero ha valore tassonomico. Attorno a questi, internamente, sono presenti numerosi microfilamenti. Occasionalmente la membrana si presenta periflagellata (estensioni pseudopodali). La cellula si presenta subsferica e/o irregolare, la stessa cosa vale per la morfologia nucleo, a volte visibile in posizione centrale, a volte in posizione basale. Il nucleolo è spesso assente. Presente il blefaroplasto. I mitocondri appaiono dispersi e i vacuoli hanno spesso inclusioni cristalline. Molto attiva la fagocitosi e la pinocitosi.
Coanociti: sono cellule flagellate fondamentali per la nutrizione delle spugne. Nelle spugne asconoidi tappezzano lo spongocele, mentre nelle siconoidi solamente i canali radiali. Formano quello che viene detto coanoderma. Il flagello è circondato da un collaretto citoplasmatico non compatto, bensì formato da un insieme di strutture più piccole addossate tra loro, che nella letteratura anglosassone sono detti microvilli o tentacoli citoplasmatici. Sono presenti 20-55 microvilli e il loro numero ha valore tassonomico. Attorno a questi, internamente, sono presenti numerosi microfilamenti. Occasionalmente la membrana si presenta periflagellata (estensioni pseudopodali). La cellula si presenta subsferica e/o irregolare, la stessa cosa vale per la morfologia nucleo, a volte visibile in posizione centrale, a volte in posizione basale. Il nucleolo è spesso assente. Presente il blefaroplasto. I mitocondri appaiono dispersi e i vacuoli hanno spesso inclusioni cristalline. Molto attiva la fagocitosi e la pinocitosi.
I flagelli non si muovono all'unisono neanche entro la stessa camera flagellata ma sono allineati in modo da spingere l'acqua nella stessa direzione.
I coanociti nelle Demospongiae e in particolare in Suberites massa possono differenziarsi in endopinacociti per regressione del flagello e dei microvilli, e in questo caso si osserva anche un’appiattimento della cellula.
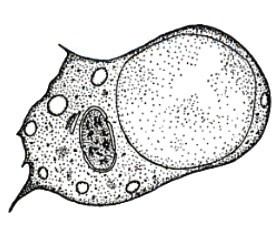
Da ricordare che i coanociti non sono sempre le cellule predominanti del coanoderma o dei canali radiali, in alcune specie sono presenti infatti in percentuale relativamente bassa. Per esempio in Haliclona elegans sono presenti solamente nel 15% delle camere. Immagine a lato.
Cellule apopilari: sono intermedie tra gli apopinacociti e i pinacociti e sono localizzate al confine tra il canale esalante e la camera flagellata. Si presentano con sezione triangolare, flagellate, e sono tipiche delle Homoscleromorpha e di alcune Demospongiae (Dendroceratida, Halisarcida e Dictyoceratida).
Archeociti: cellule totipotenti in grado di differenziarsi in altri tipi cellulari. Presentano un nucleo molto grande, sono molto mobili e contengono diversi tipi di enzimi digestivi. Possono fagocitare materiale dai canali acquiferi o accettare materiale fagocitato dai coanociti. Possono originare gli amebociti, i gametociti e anche altri pinacociti.
Amebociti: comprendono più tipi cellulari. Tra questi gli scleroblasti, gli spongioblasti, i collenciti e i miociti.
- scleroblasti (o sclerociti), sono le cellule responsabili della spicologenesi, ovvero sono le cellule che producono le spicole. Nel caso di spicole calcaree si parla di calloblasti, se silicee di silicoblasti. Sono suddivisi anche in macro- e micro- sclerociti in relazione al tipo di spicole che producono. Le spicole silice sono prodotte in un vacuolo formato dal silicalemma. Nella prima parte della spicologenesi gli sclerociti producono il filamento assiale, che contiene una bassa percentuale di silicio. Questo avviene nelle Hadromerida, Halichondrida, Spirophorida e nelle Astrophorida. Nelle Demospongiae gli sclerociti derivano dagli archeociti, mentre i megasclerociti delle spugne calcaree sembrano molto simili ai pinacociti. Al termine della spicologenesi, la cellula si dissolve.
- spongioblasti (o spongiociti), sono le cellule produttrici delle fibre di spongina perispicolare di tipo B, una proteina coinvolta nel processo di fusione delle spicole. Esistono diversi tipi di spongina, e a volte sostituisce del tutto o in parte le spicole, contribuendo da sola al sostegno della spugna. Nelle spugne silice sembrano avere chemiotassi positiva verso il siliceo. Nelle Demospongiae operano in gruppo e depongono la spongina attorno alle spicole o attorno ai grovigli di fibrille. Il citoplasma presenta un reticolo citoplasmatico molto evidente e polarizzato, e delle inclusioni dense ricche di precursori del collagene.
- collenciti, sono le cellule produttrici della spongina di tipo A. Si tratta di una varietà di fibroblasti anch’essi descritti da Sollans nel 1888. Si presentano poco mobili o fisse, stellate o fusiformi, Il citoplasma contiene delle inclusioni caratteristiche e la cellula sembra svolgere un ruolo immunitario e secretorio, spesso si può notare esternamente o adeso alla loro membrana materiale quali fibrille di collagene e di altra natura.

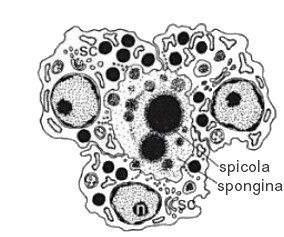
Sclerocito, a sinistra e Spongiocito a destra. Fonte: Patricia R. Bergquist - Sponges. 1979
Più amebociti possono aggregarsi e formare una gemma esterna detta sorita, per poi staccarsi dalla spugna madre e scendere sul fondale per generare un nuovo individuo.
Miociti: sono cellule proprie delle Demspongie più evolute. Regolano, in queste spugne, l'apertura dell'osculo e dei pori dermali, grazie alla loro capacità contrattile. In Microciona prolifera sono presenti due tipi di filamenti contrattili, da 50-70 nm di diametro e che formano un cluster (aggreato) nella cellula, e altre da 150-250 nm,. In Tedania ignis il diametro sale sino a 300 nm. Nelle spugne di acqua dolce mancano i miociti e sono gli endopinacociti in prossimità dell’osculo ad avere capacità contrattili. I tempi di risposta dei miociti sono lunghi e non rispondono, a differenza delle vere fibre muscolari, agli stimoli elettrici.
Lofociti: (dal greco lóphos, cresta, ciuffo e -cito) una variante del collencita. Sono cellule ameboidi che producono secreti extracellulari fibrillari. La cellula presenta una polarità. Il polo anteriore presenta numerosi pseudopodi, mentre il polo posteriore si presenta ricco di fibrille di collagene in una matrice di natura glicoproteica. Queste cellule sono molto mobili, e in alcune specie come Chondrosia sono altamente irregolari e si presentano con numerosi fillopodi che circondano la cellula. Sono presenti solo in alcune specie di spugne, come le spugne calcaree, di acque dolci e in Hadromerida.
Lofocito. Fonte: Patricia R. Bergquist - Sponges. 1979
Cellula centrale: sono cellule presenti nelle camere coanocitarie individuate da Sollas nel lontano 1888, la cui struttura è stata descritta però quasi 100 anni più tardi. La cellula è interposta tra la camera coanocitaria e la camera radiale e regola il flusso dell'acqua in ingresso. E' presente quindi nelle spugne leuconoidi.
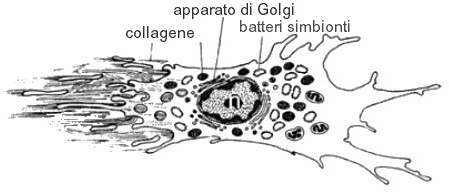
Bacteriociti: cellule ameboidi mobili contenenti procarioti simbionti. Descritti per la prima volta in Aplysina cavernicola, negli anni ’70 del secolo scorso, i batteri simbionti e le relative cellule che li ospotano sono state descritte solo nelle Demospongiae. Alcune cellule hanno un solo grande vacuolo, mentre altre numerosi e piccoli vacuoli ospitanti i batteri. Sono stati descritti anche nelle specie Petrosia ficiformis, Bemna ehrembergi e S. domuncula. Più recentemente sono stati descritti anche nella spugna carnivora Asbestopluma hypogea (Vacelet, 1996). Nella spugna C. reniformis è stato dimostrato che i batteri simbionti penetrano nella cellula nella fase embrionale (probabilmente derivano dagli archeociti) per poi mantenersi intatti per tutto lo sviluppo della cellula, sino alla metamorfosi. La trasmissione batterica è di tipo verticale. In Haliclona tubifera sono stati individuati anche nello stadio larvale di parenchimella.
Cellule vacuolari: comprendono le cellule che possiedono vacuoli che risultano essere trasparenti agli elettroni. Al microscopio elettronico risultano completamente bianchi, e in lingua anglosassone sono detti light vacuoles. Nel loro citoplasma sono sempre presenti piccoli fagosomi. Producono una sorta di idroscheletro grazie alla sintesi di collagene. Si suddividono in:
- cistenciti: si presentano con vacuoli molto grandi contenenti polisaccaridi amorfici, presenti nelle spugne di acqua dolce. Figura qui sotto, a destra (Fonte: Patricia R. Bergquist);
- Cellule globifere: con citoplasma ridotto e uno o due grandi globuli di diversa natura citologica. Il nucleo appare piccolo e irregolare, a volte conico, e occupa quasi l’intero volume cellulare, essendo scarso il citoplasma. In alcune spugne (Hymeniacidiom heliophyla) sono coinvolti nella ricostruzione allogenica;
- Cellule sferulose: poco studiate, contengono sferule di vario tipo e di contenuto da identificare. Le sferule sono facilmente distinguibili dai fagosomi. Sono localizzate soprattutto nella porzione corticale del mesoilo, per esempio in Halisarca dujardini, Crambe crambe, Crambe reniformis e Iophon piceum. Nella spugna C. reniformis le sferule contengono metaboliti tossici, mentre in Aplysina sp. e Plakina trilopha le sferule sono coinvolte nell’escrezione, in Axinella accumulano lectina. Quindi il ruolo delle sferule sembra essere decisamente eterogeneo. Interessante il fatto che alcune sferule penetrano nella larva o addirittura gia nella fase di prelarva delle spugne. Probabilmente le sferule hanno anche un ruolo immunitario e secretivo;
- Cellule granulari: sembrano essere una variante delle cellule sferulose, infatti l’unica differenza è relativa alla dimensione delle sferule. Nella specie H. dujardini le sferule contengono sostanze antibatteriche e antimicotiche (Krilova et al., 2004);
- Cellule rabddifere: larghe, ovali o fusiformi. Contengono numerose inclusioni acide a forma bastoncellare come mucopolisaccaridi che rilasciano nel mesoilo;
- Cellule con piccole inclusioni: comprendono le cellule grigie o glicociti, descritti per la prima volta in M. prolifera. Il nome deriva dalla colorazione grigia che permette di distinguerle dalle cellule circostanti. Sono coinvolte nel metabolismo del glicogeno e nella secrezione del collagene;
- Cellule microgranulari: ricche di piccoli granuli densi agli elettroni. Poco note, hanno spesso il nucleo privo di nucleolo. La loro funzione è sconosciuta. Forse coinvolte nella produzione di glicoproteine.
Cellula rabdifera, con le tipiche inclusioni a bastoncello la cui funzione rimane poco nota.
FINE PRIMA PARTE
ARTICOLI CORRELATI
- SPUGNE, NON SOLO PER FARSI IL BAGNO
- I PORIFERI Parte II Le spugne perforanti
- I PORIFERI Parte III (in preparazione)
BIBLIOGRAFIA
- Alexander V. Ereskovsky - The Comparative Embryology of Sponges. Springer 2010. Libro specialistico, aggiornato e monografico sull'embriologia delle spugne.
- Ereskovsky A. V. et al. The Homoscleromorph sponge Oscarella lobularis, a promising sponge model in evolutionary and developmental biology. Volume 31, Issue 1, pages 89–97. 2009
- Joachim Reitner, Helmut Keupp - Fossil and recent sponges. Freie Universität Berlin. Institut für Paläontologie. 1991
- Patricia R. Bergquist - Sponges. 1979. Libro dal quale abbiamo tratto le immagini delle cellule in bianco e nero, ormai introvabile, e ancora attuale.
- Pavans de Ceccatty, M - Cytoskeletal organization and tissue patterns of epithelia in the sponge Ephydatia muelleri. J. Morphol. 189,45 –65. 1986
- Thomas C. G. Bosch - The process of Archeocyte differentiation and the cell system of E. fluviatilis revealed using molecular markers. In Stem Cells: From Hydra to Man
- Tracy L. Simpson - The cell biology of sponges. 1984
- Vacelet, J. Boury-Esnault, N - A new species of carnivorous sponge (Demospongiae: Cladorhizidae) from a Mediterranean cave. Pp. 109-115. In: Willenz, Ph. (Ed.), Recent Advances in Sponge Biodiversity Inventory and Documentation. Bulletin de l’Institut royal des Sciences naturelles de Belgique. Biologie. 1996
SITOGRAFIA
Questo articolo è protetto da Copyright © e non può essere riprodotto e diffuso tramite nessun mezzo elettronico o cartaceo senza esplicita autorizzazione scritta da parte dello staff di BiologiaMarina.eu.
Ideazione: Pierfederici Giovanni - Progetto: Pierfederici Giovanni, Castronuovo Motta Nicola.
Prima Pubblicazione 31 Lug 2006 - Le immagini dei Collaboratori detentori del Copyright © sono riproducibili solo dietro specifica autorizzazione.
Si dichiara, ai sensi della legge del 7 Marzo 2001 n. 62 che questo sito non rientra nella categoria di "Informazione periodica" in quanto viene aggiornato ad intervalli non regolari
XHTML 1.0 Transitional – CSS
