
I CORALLI ERMATIPICI
I coralli ermatipici, che edificano i grandi reef che tutti conosciamo, appartengono alla classe Antozoa. Insieme alle classi Idrozoa, Scifozoa, e Cubozoa, costituiscono il Phylum CNIDARIA (knide = ortica) che comprende oltre 9000 specie diffuse nei mari di tutto il globo. Questi organismi sono caratterizzati da un tipo di simmetria detta RADIALE o RAGGIATA, anche se a volte tende a modificarsi in BIRADIALE o QUADRIRADIALE. E' possibile quindi individuare un asse di simmetria (ASSE ORO-ABORALE), attorno al quale sono disposte circolarmente tutte le parti che costituiscono l'organismo. La presenza della cavità boccale definisce la polarità dell'animale. Gli Cnidari sono caratterizzati da due distinte forme, il polipo e la medusa, che si alternano durante il ciclo vitale. L'anatomia risulta abbastanza semplice, il corpo consiste infatti in un epitelio esterno detto ECTODERMA o EPIDERMA, e di uno interno detto ENDODERMA o GASTRODERMA. Gia allo stadio larvale si distinguono ectoderma ed endoderma, e tale condizione è detta DIBLASTICA (due layers, cioè due strati). Il gastroderma delimita quindi nella forma adulta la cavità digestiva, l'unica cavità interna degli Cnidari, che svolge una duplice funzione, digestiva e circolatoria. Negli Scifozoi, Antozoi e Cubozoi tale cavità tende ad essere un pò più complessa. Nella classe Scifozoa, il cui ciclo polipo-medusa è dominato da quest'ultima, tipicamente la bocca porta ad una sorta di corto imbuto detto STOMODEO (sinonimo di FARINGE ANTERIORE o INTESTINO ANTERIORE), per poi confluire nella cavità gastrica che in molti casi presenta quattro TASCHE GASTRALI che contengono FILAMENTI GASTRICI ricchi di CNIDOBLASTI, le cellule urticanti qui con funzione digestiva. La classe Cubozoa è caratterizzata spesso da una cavità gastrica con TASCHE e SETTI GASTRALI, che sono però assenti nelle forme larvali (CUBOPOLIPI). Infine nella classe Antozoa la cavità boccale (DISCO ORALE) risulta appiattita e posta in prossimità dello STOMODEO, come negli Scifozoi, che è provvisto di muscoli e quindi può chiudersi per trattenere le prede. La cavità gastrica è poi provvista di SETTI MESENTERICI (o SARCOSETTI) che si saldano perfettamente allo stomodeo, mentre rimangono liberi nella porzione distale. Dai margini liberi dei setti si dipartono dei FILAMENTI MESENTERICI che si possono prolungare in filamenti liberi e che possono essere estroflessi attraverso l'apertura boccale o anche attraverso le pareti del corpo.
Gli Cnidari possiedono oltre ad un sistema digestivo, anche un sistema muscolare, nervoso, sensoriale (le cubomeduse hanno veri occhi detti globi oculari con tanto di cristallino, cornea e retina), e un primitivo sistema di sostegno. Mancano di un distinto sistema respiratorio, circolatorio e riproduttivo. La riproduzione può essere asessuale o sessuale. Nei polipi la riproduzione asessuale avviene per SCISSIPARITA' (divisione del corpo in due parti che ricostituiscono un polipo completo) o per GEMMIPARITA' (da escrescenze del corpo dette gemme o stoloni si origina un altro individuo che può staccarsi o meno, in quest'ultimo caso si originano delle colonie). Infine la riproduzione asessuale può avvenire per STROBILAZIONE, in tal caso per frammentazione trasversale del corpo si originano le EFIRE, a forma di stella e in grado di nutrirsi autonomamente, che poi daranno origine a meduse a sessi separati. La riproduzione sessuale passa attraverso la formazione di gameti (le cellule riproduttive) prodotti da cellule gastrodermali. Diffuso sia l'ermafroditismo (un unico individuo produce entrambi i tipi di gameti) che il gonocorismo (sessi separati e, nel caso degli cnidari, non distinguibili morfologicamente).
In questo grande gruppo dall'uovo si sviluppa una larva liberamente natante detta PLANULA, la quale può condurre vita autonoma per molto tempo, prima che avvenga la metamorfosi. In questa sede non tratteremo le tipologie larvali, precisiamo tuttavia che la planula non è l'unica forma larvale del Phylum, poiché tra gli Scifozoi e gli Idrozoi si sviluppa anche l'ACTINULA, mentre tra gli Antozoi si ha anche la LARVA ALCAMPOIDE e la LARVA di SEMPER'S, la LARVA EDWARDSIA, la CERINULA (tipica dei Ceriantari, che sono Antozoi simili ad anemoni), e infine le larve pelagiche ZOANTHINA e ZOANTHELLA (tipiche dei Zoantari, che sono gli Antozoi più primitivi), oltre a variazioni più o meno importanti sul tema che si manifestano durante la metamorfosi larvale (per esempio lo SCIFOSTOMA negli Scifozoi). Le larve dopo un periodo più o meno lungo si adagiano su un substrato idoneo (processo detto di settlement) dove possono dar vita a nuovi polipi e a nuove colonie.
ZOOLOGIA DEGLI ANTOZOI
Cominciamo ora ad esaminare più in dettaglio gli Antozoi. I coralli* che comunemente siamo abituati a vedere nelle immersioni dei mari tropicali, appartengono alla classe Antozoa (da ánthos + zôion = fiore + animale). Questa classe di organismi unicamente marini, si è evoluta perfezionandosi solamente sullo stadio di polipo, mentre la medusa manca completamente. I polipi possono essere solitari o coloniali, e sono generalmente più complessi dei loro omologhi apparteneti alle altre classi. Esaminiamo la figura sottostante:
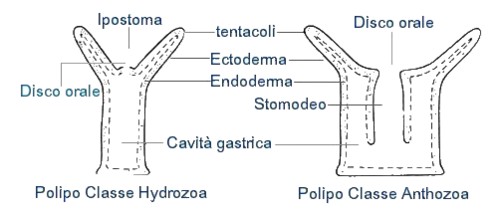
E' possibile notare che nei polipi appartenenti alla classe degli Idrozoi, la cavità gastrica è tapezzata da cellule dell'endoderma (tratteggio), mentre negli Antozoi si ha una invaginazione che costituisce lo stomodeo che è tapezzato di cellule dell'endoderma (linea non tratteggiata). Si tratta di un vantaggio evolutivo poiché essendo lo stomoideo contrattile, consente di trattenere le prede catturate, e al tempo stesso di convogliare acqua alla cavità gastrica, attraverso una doccia ciliata liscia nota con il termine di SIFONOGLIFO. Gli OTTOCORALLI o Alcionari hanno otto tentacoli e otto setti mesenterici, mentre gli ESACORALLI o Zoantari hanno sei setti mesenterici (o multipli di sei) e un gran numero di tentacoli. Le forme che hanno 12 o più setti, pervengono da larve con otto setti, ai quali se ne aggiungono altri di secondo e terzo ordine e che tuttavia non si saldano allo stomodeo, quindi sono setti mesenterici incompleti. I setti mesenterici oltre ad aumentare la superficie della cavità gastrica, sono anche la sede di formazione dei gameti. Gli Antozoi catturano le loro prede quali molluschi, crostacei, pesci e altri piccoli invertebrati attraverso i loro tentacoli urticanti, ricchi di cellule specializzate e urticanti chiamate CNIDOCITI. L'alimento è poi convogliato alla cavità gastrica dove viene assorbito dalle cellule del gastroderma e il materiale indigesto viene espulso attraverso il disco orale. Non meno importante a livello di alimentazione è il ruolo svolto dalle alghe simbionti, che illustreremo tra poco. Le specie di dimensioni modeste utilizzano il plancton, la POM, acronimo utilizzato in biologia marina per indicare il materiale organico particellare, e anche la DOM altro acronimo che indica il materiale organico disciolto, assorbito anche attraverso l'endoderma. Molte specie vivono solitarie, adese ad un substrato fisso oppure mobile, è il caso per esempio della specie Calliactis parasitica, che vive in simbiosi con il paguro Pagurus arrosor o P. oculatus, si tratta di un rapporto mutualistico e entrambe le specie ricevono dei vantaggi. Il paguro utilizza l'attinia come arma difensiva, e l'attinia sfrutta sempre nuovi ambienti grazie alla mobilità del paguro. Alcune attinie allungano attraverso la loro porzione terminale (il DISCO BASALE) l'apertura della conchiglia così il paguro crescendo non ha necessità di cambiare "abitazione". I pesci del genere Amphiprion vivono in simbiosi con le enormi attinie Stoichactis della grande barriera corallina australiana, e possono rifugiarsi tra i tentacoli urticanti senza subirne gli effetti. Alcune attinie possono anche staccarsi dal substrato e nuotare, agitando energicamente i tentacoli, è il caso di alcune forme di piccole dimensioni come Boloceroides, altre invece come Cerianthus possono scavare sulla sabbia lasciando esposti solamente i tentacoli.
I REEF CORALLINI
Ma l'aspetto che più colpisce e che più caratterizza i coralli ermatipici è la loro capacità di costruire un massiccio scheletro calcareo, grazie alla possibilità che hanno di secernere proteine fibrose associate a carbonato di calcio. In molte specie i polipi alloggiano entro una strutura a coppa, formati da cristalli di carbonato di calcio dalla caratteristica forma a ventaglio, e disposti secondo uno schema tipico di ogni specie. Lo scheletro è costituito da aragonite, che è una forma cristallina del carbonato di calcio, mentre è assente la calcite che è invece una delle forme cristalline più comuni del carbonato di calcio. Il meccanismo biochimico che sta alla base della deposizione dello scheletro è complesso è non del tutto chiaro. Nei primi anni '90 studi di microscopia hanno evidenziato che i cristalli di carbonato di calcio sono dislocati entro vescicole delle cellule dell'ectoderma, la parte esterna del corallo, e successivamente vengono espulsi all'esterno dove fungono da nuclei di condensazione per altri cristalli, in modo da permettere la crescita dello scheletro. I coralli costruttori dispongono anche dell'enzima anidrasi carbonica, che accellera la conversione della CO2 prodotta attraverso la respirazione in acido carbonico H2CO3, il quale spontaneamente si scinde poi nello ione carbonato e bicarbonato. Il fattore principale che interviene sulla crescita dello scheletro è la luce, ed è sorprendente come il tasso di crescita sia influenzato da variazioni della copertura del cielo, dalla trasparenza delle acque, dalla durata del giorno e ovviamente dalla profondità. Il tasso di crescita può essere anche di 15 volte superiore nelle ore diurne rispetto a quelle notturne, e questo perchè il ruolo delle alghe simbionti, le zooxantelle, è fondamentale. Se tali alghe vengono allontanate o espulse, il tasso di crescita si abbassa notevolmente e si mantiene pressochè invariato durante il giorno, senza significative differenze con la notte. Il tutto è spiegabile con la capacità che le zooxantelle hanno di fissare la CO2 attraverso la fotosintesi e che determina un aumento della concentrazione dello ione carbonato a livello dei polipi. Nel complesso le alghe simbionti svolgono un importante ruolo sinergico che contribuisce ad aumentare di decine di volte il tasso di crescita dei banchi corallini.
LE ZOOXANTELLE
Le zooxantelle sono normalmente alloggiate all'interno di ciascuna cellula della parete gastrica dei polipi, ma possono essere posizionate anche in altre parti, per esempio all'esterno, ma in tal caso esse non contribuiscono alla crescita dello scheletro calcareo. A volte sono assenti, tuttavia non è questo il caso dei coralli edificatori, che necessitano sempre delle loro ospiti fotosintetiche. Alcune specie di zooxantelle possono albegare in specie differenti di coralli, altre si sono specializzate per singole specie. Le alghe durante il giorno assorbono CO2, fosfati ed ammoniaca, e producono ossigeno, che essendo in eccesso è utilizzato solo in parte dai polipi. Le stesse alghe assumono durante il giorno anche i metaboliti di rifiuto prodotti dal metabolismo dei coralli, utilizzandoli per la fotosintesi e convertendoli in nuova materia organica. Le sostanze che le alghe forniscono ai polipi (amminoacidi, zuccheri e glicerina) non sembrano essere così essenziali per il loro nutrimento, i polipi sono abili nel catturare le loro prede, e quindi sono pressochè autonomi dal punto di vista alimentare. Tuttavia come detto il tasso di crescita della struttura scheletrica dei polipi diminuisce sensibilmente se vengono eliminate le zooxantelle. Perchè accade questo? Negli anni '90 il biologo marino T. J. Goreau dimostrò che circa 2/3 della CO2 prodotta dalla respirazione dei polipi finisce nel loro scheletro carbonatico. Esiste in altre parole una via interna di riutilizzo della CO2, che è sostenuta grazie alla presenza delle alghe simbionti. Senza le alghe tale quantità di carbonio inorganico andrebbe persa. Le altre fonti di carbonio, come ione carbonato, sono rappresentate dall'acqua di mare e, come carbonio organico, dagli organismi che costituiscono l'alimento per i polipi.
LA RIPRODUZIONE DEGLI ANTOZOI
Spawning è il termine utilizzato per indicare l'imponente riproduzione in massa che interessa le madrepore della grande barriera australiana. L'evento è complesso e regolato da precisi orologi biologici (fasi lunari, ciclo delle maree e temperatura dell'acqua). Qualche giorno dopo l'ultimo plenilunio di primavera è possibile, per chi ha la fortuna di trovarsi sul posto, ammirare una sorta di "nevicata al contrario" dove milioni di colorati involucri salgono verso la superficie liberando in una sorta di piccola esposione celule uovo e spermatozoi. Lo spawning interessa contemporaneamente non solo colonie della stessa specie, ma anche quelle di altre specie, così per pochi giorni all'anno si concentrano in mare una miriade di gameti che il giorno successivo colorano, galleggiando, la superficie del mare. La scoperta di questo fenomeno risale al 1981, quando si scoprì che sette specie si riproducevano nella stessa notte. Nel 1982 le specie scoperte erano gia più di 30. Tale evento sincronizzato interessa praticamente tutta la grande barriera australiana, lungo tutti i 2000 Km del grande reef. Moltissime specie sono ermafrodite, e producono circa sei mesi prima del grande evento i gameti femminili, mentre quelli maschili sono prodotti circa 2 mesi dopo. I gameti si raggruppano a formare quelle che comunemente sono chiamate gonadi, tuttavia precisiamo che non si tratta di veri organi riproduttori, trattasi più che altro di un ammasso di cellule germinali, alloggiate all'interno della cavità gastrica in prossimità dei setti mesenterici. I ricercatori della James Cook University australiana hanno previsto per molti anni la data esatta dello spawning, che avviene sempre una o due notti dopo il pleilunio, al termine della primavera o all'inizio dell'estate. Il momento esatto dipende fortemente dalla temperatura dell'acqua e quindi occorre "solo" monitorarne l'andamento per prevederne il giorno esatto. Una volta in superficie i sacculi si aprono e fuoriescono i gameti. Spermatozoi e cellule uovo si incontrano grazie a segnali biochimici e danno vita, dopo la fecondazione ad una larva planctonica liberamente natante detta PLANULA. P er alcune specie, gia dopo tre giorni la planula è pronta a fissarsi ad un nuovo substrato dal quale si originerà una nuova colonia, anche molto lontana da quella progenitrice, per altre specie invece i tempi sono più lunghi e non sono comunque sempre noti. In laboratorio alcune larve sono rimaste tali per oltre sei mesi. Quali sono i vantaggi e gli svantaggi di un evento massivo e sincronizzato come quello appena descritto? Riprodursi di notte e massivamente ha il vantaggio di limitare la predazione da parte dei pesci planctivori, che hanno per lo più abitudini diurne, e di saziare ben presto altri organismi che si nutrono di plancton garantendo la sopravvivenza di un gran numero di gameti. Inoltre l'alta densità garantisce l'incontro tra gameti femminili e maschili appartenenti alla stessa specie. Lo svantaggio riguarda la competizione per il substrato che le future larve si appresteranno a colonizzare, ma soprattutto ha a che fare con eventi naturali (o di origine antropica) che potrebbero uccidere l'intera flottiglia di gameti in un solo giorno. Per esempio anni fa le piogge torrenziali hanno determinato lo sversamento a mare da parte dei fiumi di grandi quantità di acque dolci che essendo meno dense di quelle salate, si sono posizionate sulla superficie uccidendo una percentuale altissima di gameti.
Dalla fusione dei gameti si origina come detto la planula. Tipicamente la planula metamorfosa in due stadi intermedi, la larva EDWARDSIA, che presenta otto setti e la larva ALCAMPOIDE, con 12 setti. Gli Antozoi Ceriantari hanno un distinto stadio intermedio, la CERINULA, che presena ectoderma flagellato con quattro tentacoli marginali. Ricordiamo infine le larve pelagiche ZOANTHINA e ZOANTHELLA (tipiche dei Zoantari). La prima ha una forma tipicamente ovale con lunghe cilia in prossimità del polo orale. La seconda ha una morfologia maggiormente allungata e con una banda ventrale di lunghe cilia vibratili.

IL PARADOSSO DI DARWIN
Il "paradosso delle barriere coralline" fu identificato e proposto da Darwin nel 1842. Egli notò infatti che le aree più ricche in termini di biodiversità si trovano in alcune delle acque più povere di sostanze nutrienti. Infatti per cause multifattoriali le acque tropicali sono anche le meno produtive del pianeta, a differenza di quelle temperate e fredde che hanno invece tassi di produzione primaria molto elevati. Le barriere coralline possono benissimo essere paragonate a delle oasi in mezzo ad un deserto, e basta allontanarsi di poco per rendersene conto. Ma come fanno le barriere coralline a sostenersi dal punto di vista biologico? Risposte esatte non ce ne sono, solo ipotesi. Tuttavia nel 2001 alcuni ricercatori del'Università di Brema hanno scoperto nelle barriere coralline del Mar Rosso una miriade di nuovi organismi che vivono all'interno delle crepe della barriera e fino ad allora mai catalogati. Si è evidenziato che la superficie disponibile all'interno delle fessurazioni sono molto maggiori rispetto a quelle disponibili all'esterno della barriera. Questi studi potrebbero quindi contribuire a risolvere il paradosso delle barriere coralline ipotizzato dal grande naturalista più di un secolo fa. Teoricamente la trattazione dei coralli ermatipici non finisce qui. Molto ci sarebbe da aggiungere, soprattutto per cio che concerne l'architettura dei reef, la loro morfologia e i fattori che la influenzano, la biogeografia, l'ecologia, la biodiversità, il fenomeno dello sbiancamento dei coralli e dell'acidificazione dei mari. Volutamente sono stati tralasciati i gruppi di Antozoi non costruttori, ma ne riparleremo prossimamente. La bibliografia sull'argomento è vastissima. Su alcuni argomenti sono in preparazione altri articoli, ma per chi volesse da subito approfondirlo consigliamo i testi riportati qui sotto in biblioigrafia.
* Il il termine "corallo" di norma si utilizza solamente quando ci si riferisce al genere Corallium. Tuttavia per motivi pratici in questa sede utilizziamo il termine "Corallo" per indicare anche altri organismi, come Madreporari e altri Antozoi.
DOMANDE (ricevuta il--/--/2009)
D:Buongiorno, leggendo l'articolo sui coralli ho notato subito l'errore su cui spesso cadete anche voi biologi marini. Il termine "coralli ermatipici" indica i coralli che sono in simbiosi con le zoxantelle, non necessariamente indica un corallo esclusivamente costruttore. Ci tenevo alla precisazione. Gaetano&Monai
R: I termini "ermatipico" e "aermatipico" sono stati introdotti da Veron nel 1986, che intervenne su una precedente definizione di Wells che per primo introdusse il termine "coralli ermatipici" nel lontano 1933, proprio per descrivere i coralli edificatori dei grandi reef ospitanti le zooxantelle. Al contario con il termine "aermatipico" indicò i coralli non costruttori e privi di zooxantelle. Con il termine "zooxantellati" invece si indicano le associazioni simbionti con zooxantelle, per cui i due termini (ermatipico e zoxantellato) divennero ben presto sinonimi. Per far luce sulla questione intervennero Schuhumacher e Zibrowius (1985) che precisarono il fatto che non necessariamente i coralli ermatipici ospitano zooxantelle, e che non necessariamente i coralli zooxantellati sono costruttori di reef. Attualmente quindi con il termine " ermatipico" si indicano solo i coralli e altri organismi capaci di edificare un reef, dove per reef si intendono strutture topografiche e biogeniche positive, capaci di resistere all'azione del moto ondoso e persistenti per un lungo periodo di tempo (Guilcher 1988). Quindi il significato non è associato alla presenza o meno delle zooxantelle, anche se è vero che la maggioranza dei coralli ermatipici sono anche zooxantellati. Così come è vero che la maggioranza dei coralli aermatipici sono anche azooxantellati. Esempi di coralli azooxantellati ma ermatipici sono stati trovati a 500 m di profondità, in assenza di luce nonché anche in acque molto fredde.
AGGIUNGI UN COMMENTO [Se non desideri visualizzare il tuo commento sul tuo profilo Facebook, ricordati di togliere la spunta della ceckbox]
BIBLIOGRAFIA
- Marine Biodiversity - Cambridge University Press 2005. Cap 9, 10
- Marine Community Ecology - Sinauer Associates Incorporated 2001
- Coral Reef Fishes - Princeton University Press 2002
- Coral Reef Conservation - Cambridge University Press 2006
ARTICOLI CORRELATI
- IL CORALLO ROSSO
- LE GORGONIE ALLA PORTA DEL MEDITERRANEO
- ANTICHI COSTRUTTORI DI SCOGLIERE CORALLINE
- MUORE IL CORALLO DELLE ISOLE ANDAMAN
Questo articolo è protetto da Copyright © e non può essere riprodotto e diffuso tramite nessun mezzo elettronico o cartaceo senza esplicita autorizzazione scritta da parte dello staff di BiologiaMarina.eu.
Ideazione: Pierfederici Giovanni - Progetto: Pierfederici Giovanni, Castronuovo Motta Nicola.
Prima Pubblicazione 31 Lug 2006 - Le immagini dei Collaboratori detentori del Copyright © sono riproducibili solo dietro specifica autorizzazione.
Si dichiara, ai sensi della legge del 7 Marzo 2001 n. 62 che questo sito non rientra nella categoria di "Informazione periodica" in quanto viene aggiornato ad intervalli non regolari
XHTML 1.0 Transitional – CSS
