
IL GIACIMENTO PALEONTOLOGICO DELLA RASA DI VARESE: UNA BARRIERA CORALLINA DI 240 MILIONI DI ANNI FA
| ABSTRACT | ||
| Rasa is an outlying district of Varese (Lombardy-Italy). Here hundreds of fossils have been found in the rock named "Rasa Dolomite" (Dolomia della Rasa, sensu Airaghi 1935): a local facies of the geological formation known as "Salvatore Dolomite", dating approximately from the Middle Triassic Period (Mesozoic Era). Paleontological research in this area began around the end of the 19th Century, and the first documented discovery was a badly preserved imprint of an ammonoid found by the geologist, T. Taramelli. He immediately took the find to E. von Mojsisovics, who mentioned it in an article in 1880 and estimated a possible date: the "Rasa Dolomite" was thus attributed to the Middle Triassic Period. In 1885, A. Tommasi published the first work on the fossil fauna of Rasa, but the examples he described were incomplete and their provenance uncertain. In 1935, C. Airaghi published a note based on several shell fossils found during excavations at the "Villa Cagnola" and the "Cava Lambertoni", close to the village of Rasa. Airaghi attributed the "Rasa Dolomite" to the Ladinian stage, thereby summarily confirming the observations made by Mojsisovics. However, the brief article by Airaghi contains no illustrations and today is outdated. Thanks to research carried out in several of the outcrops situated near the village of Rasa (excavations during building work or road maintenance), it was possible to examine a remarkable amount of material, which highlights a highly varied fauna with approximately a hundred species of marine invertebrates, as well as the remains of marine and land vertebrates. Because of the remarkable variety of ammonoids, dating has finally been established as between the end of the Upper Anisian and the Lower Ladinian stage, with a greater wealth of fauna in the strata identified as the Anisian/Ladinian boundary. This allows a comparison between the deposit at Rasa and the more wellknown one at Besano/Monte San Giorgio (UNESCO 2003-2010), which is almost contemporary but settled in a different paleoenvironment (lagoon). The layering of the "Rasa Dolomite" clearly occurred in a well-oxygenated sea (coral reef), with large deposits of shells that form the hills of Monte Chiusarella. These sediments appear to have accumulated all around the permian volcano that emerged from the sea, now known as Monte Martica and situated between Rasa and the village of Brinzio. The individual species of sea molluscs found in the “Rasa Dolomite”, show a certain affinity with the fauna of the Ladinian stage of the Lombard Prealps and the Dolomites, and the fauna of the Muschelkalk in Silesia (Poland), especially as regards the more primitive gasteropods (such as Kittlidiscus lottneri=Euomphalus lottneri) and certain bivalves (e.g. Palaeolima acutecostata). Particularly interesting are several nautiloids (Mojsvaroceras kummeli), typical of the Negev in southern Israel, and the gasteropod Scurriopsis cingulata, endemic of Rasa. For the first time, vertebrate remains in the Salvatore Dolomite have been studied: the teeth of the shark Asteracanthus and of the reptiles Helveticosaurus and Ticinosuchus. All the material described is housed in the "A. Stoppani" Museum of Venegono Inferiore in the province of Varese. | ||
La Rasa di Varese è una frazione del comune di Varese (fino al 1928 era frazione di Velate), ad est del Sacro Monte, situata nella Valle della Rasa, tra il M. Legnone e il M.Chiusarella, dove si trova una delle sorgenti del fiume Olona. Il paese ha un'altitudine compresa tra i 520 e i 600 m sul livello del mare. Le rocce affioranti nella zona della Rasa di Varese (VA), sono costituite da dolomie calcaree, ovvero la "Dolomia della Rasa" (Airaghi, 1935): una facies locale della formazione geologica detta "Dolomia del S. Salvatore", databile per lo più al Triassico Medio (Era Mesozoica).

Sopra, panorama della Rasa di Varese.
Grazie ad una serie di ricerche in vari affioramenti situati proprio in corrispondenza dell'abitato della Rasa (scavi a scopo edilizio o per manutenzione stradale), è stata esaminata una notevole quantità di materiale, che ha messo in evidenza una fauna ben diversificata, con circa un centinaio di specie di invertebrati marini, oltre alla presenza, mai documentata prima in questa formazione sedimentaria, di resti di vertebrati marini e terrestri. La datazione, chiarita solo ora con precisione dalla notevole varietà degli ammonoidi, viene delimitata tra la parte finale dell’Anisico Superiore e il Ladinico Inferiore, con la maggior ricchezza faunistica negli strati identificati come limite Anisico/Ladinico. Ciò permette di paragonare il giacimento della Rasa a quello di Besano/Monte S. Giorgio (UNESCO 2003, 2010), che risulta pressochè contemporaneo, ma con deposizione in ambiente diverso (lagunare). La deposizione della "Dolomia della Rasa" è invece chiaramente avvenuta in ambiente di mare ben ossigenato (barriera corallina), con grandi depositi di gusci di conchiglie, che hanno formato le alture del Monte Chiusarella. Tali sedimenti sembrano essersi accumulati tutt'attorno ad un vulcano del Permiano che costituiva una zona emersa dal mare: l’attuale Monte Martica, situato tra la Rasa e il paese di Brinzio.
Per quanto riguarda le singole specie di molluschi marini rinvenute nella "Dolomia della Rasa" e qui descritte, si è potuta osservare una certa affinità sia con le faune del Ladinico delle Alpi e Prealpi lombarde e della zona dolomitica, sia con la fauna della dolomia del Muschelkalk della Slesia (Polonia), soprattutto per i gasteropodi più primitivi (come Kittlidiscus lottneri) e per alcuni bivalvi (come Palaeolima acutecostata). Di particolare interesse, sono poi alcuni nautiloidi riferibili alla specie Mojsvaroceras kummeli, tipica e quasi esclusiva dei giacimenti triassici del Negev in Israele, e ora segnalata per la prima volta in Italia. Da citare è anche il ritrovamento di Scurriopsis cingulata che si ritiene esclusiva della Rasa.
PALEOGEOGRAFIA E PALEOAMBIENTE
Alla fine dell'era Paleozoica la Pangea si era fratturata in due supercontinenti: la Laurasia a N e il Gondwana a S. La zona che corrisponde grosso modo alla nostra penisola, che ancora non esisteva, si trovava nella parte occidentale dell'oceano chiamato Tetide. Nel Triassico medio l'area corrispondente alle Prealpi odierne era caratterizzata da un ambiente simile a quello che oggi si trova nel Golfo Arabico. Isole e vulcani permiani emergevano da un mare poco profondo, nel quale si stavano formando scogliere biocostruite principalmente da alghe calcaree diplopore. Ampie zone di mare basso con fondali piatti, costituiti da accumuli di resti di organismi (piattaforme carbonatiche), si estendevano attorno alle terre emerse. A questi erano associate zone asfittiche là dove la limitata circolazione di acqua sul fondo quasi stagnante, non consentiva agli organismi bentonici di sopravvivere. Da questi bacini hanno origine gli scisti bituminosi di Besano e di Monte S. Giorgio.
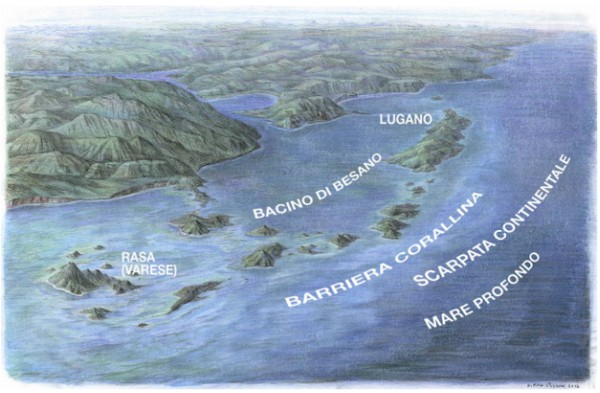
Per questo motivo, abbiamo nello stesso lasso di tempo, a Besano, una deposizione ben stratificata, con resti di vertebrati conservati in connessione anatomica, ed altri organismi che comunque non vivevano sul fondo marino (necton). Mentre alla Rasa, come al Monte S. Salvatore di Lugano (da cui prende nome la "Dolomia del S. Salvatore") la deposizione è stratificata in grandi banchi dolomitici. Tale sedimentazione è poco favorevole alla conservazione di vertebrati in connessione anatomica, mentre è ricca di una notevole varietà di molluschi ed altri organismi bentonici, come mitili, coralli, ricci di mare, gasteropodi ecc. Oltre agli organismi bentonici, si accumulavano gusci di nautiloidi e ammonoidi (necton) del tipo trachiostraca (cioè con scultura della conchiglia ben rilevata), adatti appunto alla vita di scogliera.

LA BARRIERA CORALLINA DELLA RASA: UN ECOSISTEMA MARINO SUBTROPICALE DI 240 MILIONI DI ANNI FA
La ricchezza faunistica della barriera corallina della Rasa, è ben riconoscibile in particolare nella parte alta del borgo (Corte del Cassinot), dove, in corrispondenza e al di sotto di un livello ricco di gigli di mare (Encrinus cf. cassianus) con coralli coloniali associati a spugne e alghe (bioerme?), sono state trovate conchiglie fossili di molluschi gasteropodi, lamellibranchi, ammonoidi, nautiloidi, echinodermi, brachiopodi, alghe calcaree e rari resti di vertebrati marini. La profondità doveva essere compresa tra il piano mesolitorale e quello infralitorale, dove la luce del sole riesce a raggiungere bene il fondale. Ciò è deducibile dalla presenza delle forme che si possono ritenere almeno in parte autoctone: alghe calcaree, coralli e spugne, gigli di mare con l’esoscheletro in buona parte in connessione anatomica, in posizione verticale rispetto alla deposizione dei sedimenti.
I vertebrati, tra l'altro si tratta di specie molto rare, che popolavano questi fondali e che sono testimoniati da ritrovamenti di denti e di frammenti di ossa, sono squali di medie dimensioni (genere Asteracanthus) con denti arrotondati, adatti a schiacciare i gusci dei molluschi di cui si cibavano, e rettili arcaici che potevano raggiungere i 3 metri di lunghezza, riferibili al genere Helveticosaurus (lo scheletro più completo fu trovato nel 1935, sul M. S. Giorgio, ma l’interpretazione dei resti di questo animale risulta ancora oggi problematica). La presenza di terre emerse nelle vicinanze è testimoniata dal ritrovamento di una porzione di dente attribuibile al rettile terrestre Ticinosuchus, un grosso predatore (l'unico esemplare integro conosciuto misura circa 2,5 metri e proviene anch'esso dal Monte San Giorgio).
Nel suo insieme la fauna della Dolomia della Rasa è davvero un esempio singolare in Italia, sia per varietà e tipologia delle specie, sia per la conservazione in dolomia, che solitamente contiene fossili mal conservati, mentre qui presenta in alcuni casi reperti di ottima qualità (pseudogusci con le strie di accrescimento e ossa e denti con le strutture interne perfettamente riconoscibili). In definitiva si tratta di un paleoambiente marino ben rappresentato ed eterogeneo, che contribuisce alla ricostruzione di quell'ecosistema che ha caratterizzato il Triassico Medio prealpino lombardo, oggi ormai di riferimento mondiale per quest'epoca. Una zona di mare basso con isole vulcaniche, atolli e barriere coralline ricche di vita, animali di un periodo di transizione: comparsi come una nuova esplosione della vita sulla Terra dopo la grande estinzione di massa della fine dell'Era Paleozoica, e anticipatori delle forme che domineranno il nostro pianeta nel Giurassico e nel Cretacico.
Si è ritenuto utile riportare brevemente l’elenco delle specie descritte nella pubblicazione del 2011: "La Rasa di Varese e i suoi fossili" di Pieroni V. (Pietro Macchione Editore), a cui si rimanda per eventuali approfondimenti. (vedi anche Pieroni V., "Fossili nel Varesotto: i reperti della Rasa di Varese". Rivista "Lombardia Nord-Ovest" Anno LXXVII, N° 3, Settembre/Dicembre 2004. FORMAS Varese.
Tale elenco, viene qui proposto in forma aggiornata alle nuove ricerche.
Elenco dei taxa divisi secondo le categorie ecologiche: bentos e necton
Flora bentonica (fitobenthos): due specie di alghe calcaree, determinate come Teutloporella herculea [STOPPANI, 1857] PIA, 1912 e Diplopora annulata [SCHAFHÄUTL, 1853] 1863.
Fauna bentonica (zoobenthos):
- Poriferi
Amblysiphonella cfr. strobiliformis Dieci, Antonacci & Zardini, 1968 e Cystothalamia sp.
- Coralli
Koilocoenia sp. e un genere non determinato della Famiglia ?Montlivaltiidae, Dietrich, 1926.
- Brachiopodi
Volirhynchia cfr. vivida (Bittner, 1890).
Tetractinella trigonella (Schlotheim, 1820).
Mentzelia cfr. mentzeli (Dunker, 1851).
Punctospirella fragilis (Schlotheim, 1814).
Lobothyris praepunctata (Bittner, 1890).
Coenothyris cfr. vulgaris (Schlotheim, 1820).
- Gasteropodi
Worthenia cfr. supraornata Kittl, 1894.
Kittlidiscus lottneri (= Euomphalus lottneri Eck, 1865)
Coelocentrus cf. horridus (Assmann, 1924)
Trachybembyx junonis (Kittl, 1894)
Trachybembyx sp.
Pleurotomaride indeterminato.
Scurriopsis (Scurriopsis) cingulata (Tommasi, 1885).
Naticopsis sp.
?Dicosmos (Hologyra) sp.
Planospirina esinensis [Stoppani, 1858-60] Kittl, 1899.
Trachynerita cfr. quadrata (Stoppani, 1858).
Trachynerita depressa (Hoernes, 1856).
Trachynerita nodifera (Kittl, 1894).
Kittliconcha cfr. alta (Assmann, 1937).
Anoptychia sp.
Hypsipleura cfr. subnodosa Klipstein in Kittl, 1894.
Telleria cfr. umbilicata Kittl, 1894.
Coelochrysalis sp.
Coelostylina sp.
Omphaloptycha pachygaster Kittl, 1894.
?Omphaloptycha cfr. turris (Stoppani, 1857).
?Omphaloptycha sp.
?Undularia sp.
Spirochrysalis sp.
- Bivalvi
Parallelodon cfr. esinensis (Stoppani, 1858).
Macrodontella cfr. lamellosa (Assmann, 1915).
Modiolus salzstettensis Hohenstein, 1913.
Protopis sp.
?Aviculomyalina undulata Zorn, 1971.
Pteria hallensis (Wöhrmann, 1889).
Bakevellia costata ticinensis Zorn, 1971.
Bakevellia cfr. costata costata Zorn, 1971.
Bakevellia (Neobakevellia) cfr. constricta (Stoppani, 1857).
Cassianella sp.
Ornithopecten sp.
?Pseudomonotis cfr. tarnowitzensis (Assmann, 1915)
Daonella fascicostata Rieber, 1969.
?Radulonectites sp. forma “a”.
?Radulonectites sp. forma “b”.
Enantiostreon scabiosum Giebel, 1856.
?Enantiostreon sp.
Newaagia cassiana (Muenster 1841).
Newaagia noetlingi Frech, 1912.
Placunopsis plana Giebel, 1856.
Mysidioptera cfr. cainalloi (Stoppani, 1858).
Palaeolima acutecostata (Assmann, 1937).
Lyriomyophoria sp.
Schafhaeutlia cfr. silesiaca (Assmann, 1937).
Unionites cfr. albertii (Assmann, 1915).
-Echinodermi
-Crinoidi
Encrinus cfr. cassianus (Laube, 1864)
Encrinus cfr. granulosus (Muenster, 1841)
-Echinidi
Serpianotiaris sp.
Fauna nectonica:
- Nautiloidi
Michelinoceras campanile (Mojsisovics, 1882).
Mojsvaroceras kummeli (Parnes, 1986).
Syringonautilus cfr. lilianus (Mojsisovics, 1882).
- Ammonoidi
Ticinites ticinensis Rieber, 1973.
"Celtites" cfr. fumagallii (Stabile, 1860).
"Celtites" cfr. taramellii Airaghi, 1912.
"Celtites" cfr. paronai Airaghi, 1912.
Aplococeras cfr. misanii (Mojsisovics, 1882).
Serpianites curionii Rieber, 1973
Serpianites airaghii Rieber, 1973.
Serpianites serpianensis (Airaghi, 1912).
Serpianites zinae (Airaghi, 1912).
Parakellnerites frauenfelderi frauenfelderi Rieber, 1973.
Parakellnerites frauenfelderi ?angustus Rieber, 1973.
Parakellnerites cfr. frauenfelderi Rieber, 1973.
Parakellnerites meriani Rieber, 1973.
Parakellnerites cfr. rothpletzi (Salomon, 1895).
Parakellnerites carinatus Rieber, 1973.
Parakellnerites sp.
Stoppaniceras variabilis Rieber, 1973.
Stoppaniceras cfr. artinii (Airaghi, 1912).
Stoppaniceras grandinodus Rieber, 1973.
Repossia acutenodosa Rieber, 1973.
Nevadites ambrosionii (Airaghi, 1912).
Nevadites dealessandrii (Airaghi, 1912).
Chieseiceras chiesense (Mojsisovics, 1882).
Praepinacoceras sp.
Flexoptychites acutus (Mojsisovics, 1882).
Proarcestes sp.
Protrachyceras cfr. archelaus (Laube, 1868).
Falsanolcites cfr. gortanii (Pisa, 1966).
Protrachyceras sp.
Due forme non ben determinabili appartenenti alla Sottoclasse Coleoidea, Bather, 1888. Alcuni resti di grossi vertebrati, attribuiti ai generi Asteracanthus, Helveticosaurus e Ticinosuchus.
Tutto il materiale paleontologico qui citato, è regolarmente denunciato alla Soprintendenza per i Beni Archeologici della Lombardia, e depositato presso il Museo A. Stoppani del Seminario Arcivescovile di Milano Pio XI, Via Pio XI 32, I-21040 Venegono Inferiore (VA).
Vittorio Pieroni, 2012
AGGIUNGI UN COMMENTO [Se non desideri visualizzare il tuo commento sul tuo profilo Facebook, ricordati di togliere la spunta della ceckbox]
Questo articolo è protetto da Copyright © e non può essere riprodotto e diffuso tramite nessun mezzo elettronico o cartaceo senza esplicita autorizzazione scritta da parte dello staff di BiologiaMarina.eu.
Ideazione: Pierfederici Giovanni - Progetto: Pierfederici Giovanni, Castronuovo Motta Nicola.
Prima Pubblicazione 31 Lug 2006 - Le immagini dei Collaboratori detentori del Copyright © sono riproducibili solo dietro specifica autorizzazione.
Si dichiara, ai sensi della legge del 7 Marzo 2001 n. 62 che questo sito non rientra nella categoria di "Informazione periodica" in quanto viene aggiornato ad intervalli non regolari
XHTML 1.0 Transitional – CSS
