
L'EVOLUZIONE DEI MOLLUSCHI BIVALVI [parte 02]
Pubblichiamo la seconda parte della tesina di Piva Lavinia, dedicata all'evoluzione dei molluschi bivalvi. (I primi tre paragrafi si trovano nel precedente articolo).
04. CURIOSITÀ: SPIRALE AUREA E LUNGHEZZE D'ONDA SOTTO LA SUPERFICIE DELL’ACQUA
Come la matematica e la fisica influenzano l’evoluzione degli organismi viventi
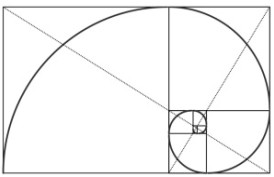
Spirale aurea
La sezione del guscio del Nautilus (Cephalopoda) è l'espressione della matematicità della spirale aurea in natura.
Attraverso la rappresentazione grafica di più rettangoli aurei, si giunge alla raffigurazione di ciò che si chiama spirale aurea.
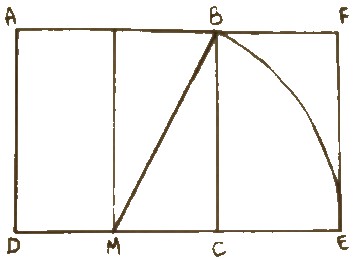
Per capire meglio come si disegna il rettangolo aureo, si parte da una base a forma quadrata di vertici A B C D. Si divide poi in due parti uguali il quadrato, con una linea verticale che passa tra i punti medi dei segmenti AB e DC. Il punto medio del segmento DC è chiamato M.
Con l'ausilio di un compasso si punta in M e lo si apre fino ad arrivare al vertice B del quadrato.
Si traccia poi un arco esteso fino a intersecare il prolungamento del segmento DC in E.
Si disegna poi il segmento EF perpendicolare a DC, e il segmento BE perpendicolare a CB.
Il rettangolo ADEF risultante è un rettangolo aureo nel quale il punto B divide il segmento AF nella sezione aurea.
Se a ogni rettangolo aureo si aggiunge sul lato maggiore un quadrato, si arriva a ottenere un altro rettangolo aureo.
Procedendo successivamente con le stesse modalità descritte sopra per il disegno dell’arco si ottiene una spirale aurea, cioè una spirale logaritmica con fattori di accrescimenti pari alla sezione aurea.
Lunghezze d’onda
Lo spettro visibile è una parte dello spettro elettromagnetico che cade tra il rosso e il violetto, e che racchiude tutti i colori percepibili dall'occhio umano. Noi percepiamo la porzione dello spettro elettromagnetico definito luce visibile.
La lunghezza d'onda della luce visibile va dai 750 nm (maggiore lunghezza d'onda, colore rosso) ai 380 nm (minore lunghezza d'onda, colore viola).
Lunghezze d'onda in mezzi come l'acqua diminuiscono in proporzione all'indice di rifrazione (grandezza che quantifica la diminuzione della velocità di propagazione della radiazione elettromagnetica, quando essa attraversa un determinato materiale).
Quando la luce incontra la superficie del mare, una certa quantità viene riflessa secondo un determinato angolo di incidenza, mentre quella che penetra all'interno, modifica la sua intensità luminosa e la sua composizione spettrale.
L'acqua assorbe in modo diverso differenti lunghezze d'onda che compongono lo spettro del visibile; in questo caso le colorazioni delle conchiglie e di molti esseri viventi che popolano il mare possono essere osservate solo in superficie, dove arriva tutta la banda dello spettro. Ad esempio, il colore rosso di molte conchiglie può essere visibile solo a bassa profondità dalla superficie dell'acqua, proprio perché questo presenta una maggiore lunghezza d'onda. Ciò si può notare dalla figura sotto. Man mano che si scende in profondità la colorazione viene influenzata dall’assorbimento delle diverse lunghezze d'onda.
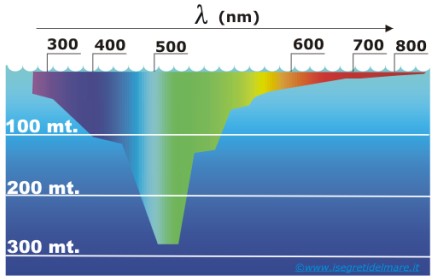
Molti pesci dai colori sgargianti e molti gasteropodi come ad esempio il Clanculus pharaonius (Linnaeus, 1758) vivono in zone superficiali del mare; così facendo le loro colorazioni risultano ben visibili e differenziano le varie specie. Leggi anche l'articolo LA RADIAZIONE SOLARE IN ACQUA.

Clanculus pharaonius (Linnaeus, 1758)
05. FILOGENESI ED EVOLUZIONE DEI BIVALVI
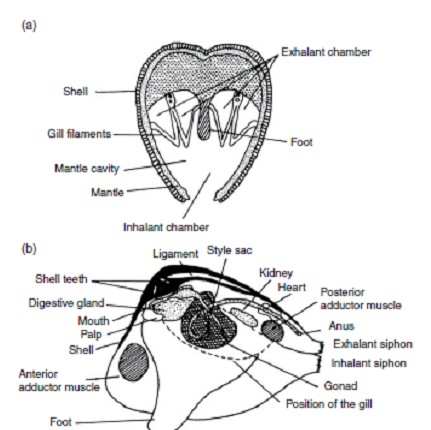
I bivalvi, Classe Bivalvia, molluschi detti un tempo anche lamellibranchi o acefali, nomi che si riferiscono rispettivamente alla conformazione delle branchie ed alla mancanza di una regione cefalica distinta, comprendono tutti i molluschi dal corpo a simmetria bilaterale, compresso e con conchiglia a due valve. La radula (parte dell'apparato boccale) è mancante, e il mantello forma due lembi che avvolgono tutto il corpo dell'animale e che si possono saldare tra loro lungo il margine ventrale, lasciando però tre aperture, una anteriore da cui sporge il piede e due posteriori per la circolazione dell'acqua nella cavità palleale. In alcune specie queste due aperture sono allungate a tubo e costituiscono i cosiddetti sifoni. Il piede può essere molto sviluppato oppure essere molto ridotto o completamente assente come nelle ostriche, che si saldano con una valva al substrato. Il piede viene usato per scavare oppure, come nel Cardium, anche per fare piccoli salti.
Nei generi Mytilus e Pinna il piede è provvisto di ghiandole del bisso, che producono i filamenti che servono per fissarsi al substrato. Alcuni, come i Lithophaga lithophaga (Linnaeus, 1758) detti datteri di mare, si insediano all'interno di rocce calcaree mediante delle secrezioni acide prodotte da apposite ghiandole. L'epitelio del mantello è deputato alla formazione della conchiglia, che racchiude il corpo dell'animale. Questa, talora, può essere ridotta o presentare irregolarità di simmetria fra le due valve, in relazione al modo di vita di certe specie, che giacciono su una valva (Pecten). I Pecten sono anche in grado di nuotare brevemente sbattendo le valve.
Le due valve sono unite dorsalmente da un legamento che le tiene aperte e la chiusura della conchiglia è dovuta alla contrazione di uno o due muscoli adduttori inseriti sulla faccia interna delle valve.
Esse sono unite dorsalmente anche da una serie di dentelli che si incastrano gli uni con gli altri: a questa formazione viene dato il nome di cerniera Il numero e la disposizione dei dentelli, così come il numero, la disposizione e lo sviluppo dei muscoli adduttori, sono elementi utili per la classificazione.
La conchiglia ha una struttura abbastanza complessa ed è fatta di tre strati: uno esterno, formato da una sostanza organica (conchiolina) detta periostraco, uno intermedio calcareo a struttura prismatica e uno interno, pure calcareo, a struttura lamellare simile all'aragonite. I tre strati sono saldati insieme e talvolta hanno uno spessore notevole; l'apice di ogni valva è detto umbone. Le branchie assumono un aspetto lamellare (da cui il nome lamellibranchi) e le lamine che le costituiscono si ripiegano così da formare quattro foglietti, che si ritrovano poi diversamente saldati nei vari gruppi.
Il tubo digerente è distinto in esofago, stomaco e intestino, che sbocca nella cavità palleale.
L'apparato escretore è formato da nefridi che si aprono con il nefrostoma nella cavità pericardica.
Il cuore è formato da un ventricolo e da due atri: il sangue arterioso dalle branchie arriva agli atri e da questi passa al ventricolo, quindi, per mezzo di arterie, si porta agli organi; esistono poi delle vene che dagli organi riportano il sangue alle branchie.
Il sangue contiene emocianina (proteina con due atomi di rame, da cui il colore verde bluastro), e talora anche emoglobina. Gli organi di senso sono organi tattili disposti in particolare lungo il margine del mantello, dove vi sono anche gli organi visivi, rappresentati da semplici cellule, oppure da organi più complessi (macchie oculari), talvolta forniti di elementi lenticolari, che sono sensibili alle variazioni di intensità della luce, ma che probabilmente non formano immagini.
I bivalvi sono nella maggior parte dei casi a sessi separati, ma talvolta sono ermafroditi con maturazione successiva dei due sessi in uno stesso individuo. In rari casi si ha un dimorfismo sessuale a carico della conchiglia che, come negli Unionidi (cozze di acqua dolce), è più grossa e rigonfia nella femmina.
La fecondazione avviene nella cavità del mantello e lo sviluppo è indiretto, con la formazione di una larva (veliger) che agli inizi conduce vita pelagica e poi diventa bentonica.
I bivalvi sono tutti acquatici e si nutrono in genere di detriti presenti sia sul fondo, sia in sospensione, che attraggono con movimenti delle ciglia presenti sull'epitelio branchiale e sulla parete interna del mantello.
La presenza di una resistente conchiglia calcarea ha fatto sì che i bivalvi si siano conservati assai bene allo stato fossile. Le loro conchiglie si rinvengono, a volte in grande abbondanza, nei sedimenti marini e d'acqua dolce di tutte le ere geologiche.
Quando non erano ancora in uso i metodi di analisi molecolari e genetici e ci si basava solo sull'analisi morfologica, si pensava che i bivalvi derivassero dai Rostroconchia, gruppo fossile estinto con la conchiglia singola allo stadio larvale e pseudo-bivalve da adulti.
I membri della classe Rostroconchia sono vissuti tra il Cambriano e il tardo Permiano, e inizialmente furono collocati tra i bivalvi, ma in seguito vennero assegnati ad una classe indipendente. Erano dotati di un guscio singolo pseudobivalve che racchiudeva il mantello e il piede muscolare. La parte anteriore del guscio probabilmente era rivolta verso il basso e aveva una fessura dalla quale emergeva il piede.
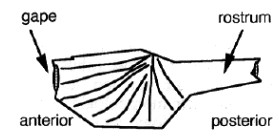
Le parti principali della conchiglia di Rostroconchia (McRoberts, 1998)
I Rostroconchia si sono quasi certamente evoluti dai molluschi Helcionelloida (Pojeta & Runnegar, 1976; Peel, 1991) durante il primo Cambriano. Inizialmente, si riteneva che essi (Runnegar & Pojeta, 1974; Pojeta & Runnegar, 1976) avessero dato luogo ai Bivalvia da un lato (per separazione delle valve e sviluppo di una adeguata cerniera), e agli Scaphopoda dall'altro (con le valve che formano un lungo tubo, e sono noti come conchiglie a zanna).
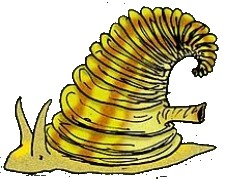
Rappresentazione di Yochelcionella cyrano (Runegar & Pojeta, 1974)
Tuttavia, successive analisi cladistiche e molecolari (Waller, 1998; Steiner & Dreyer, 2002) hanno accertato che i bivalvi siano solo lontani parenti degli scafopodi, mentre i cefalopodi (polpi, seppie, totani, calamari, Nautilus ecc..) sarebbero strettamente correlati agli scafopodi.
Le analisi embriologiche portano a sostenere la tesi che gli scafopodi siano evoluti dai Rostroconchia (Engeser & Riedel, 1997). Inoltre i cefalopodi (Pojeta, 1980) o forse sia scafopodi che cefalopodi (Runnegar, 2002; Steiner & Dreyer, 2002), sarebbero strettamente correlati, o discendenti dagli Helcionelloida. Questi Helcionelloida primitivi possono essere visti come gli antenati dei Rostroconchia (Wagner, 2011) La conseguenza potrebbe essere che gli Helcionelloida avrebbero dato luogo sia ai Rostroconchia, che ai cefalopodi, ma non ai bivalvi che, come abbiamo visto precedentemente, sembrano invece avere un progenitore comune coi gasteropodi.
Nonostante il fatto che i bivalvi, a causa dei loro gusci duri e resistenti siano gli animali che hanno lasciato la maggiore documentazione fossile, la loro sistematica, fino a tempi recenti, è stata piuttosto controversa.
Questa situazione è cambiata quando i ricercatori hanno iniziato ad applicare analisi filogenetiche su larga scala, utilizzando combinazioni di fonti di dati morfologici, paleontologici e molecolari (Bieler & Mikkelsen 2006).
Storicamente, c'è stata una pesante dipendenza da analisi che prendevano in esame singoli caratteri, ad esempio la cerniera della conchiglia, i suoi denti, la struttura delle branchie, la morfologia dello stomaco, del mantello ecc...
La situazione è cambiata nel 1970 con lo sviluppo di una sistematica numerica basata sull'analisi simultanea di più sistemi di caratteri. Dai primi anni '90 del secolo scorso i dati di sequenze geniche si sono resi disponibili, e nel corso degli ultimi due decenni queste fonti di dati hanno dato un contributo significativo agli studi sistematici che comprendono tutti i bivalvi (Giribet 2008; Plazzi & Passamonti 2010; Plazzi at al. 2011; Tsubaki et al., 2011; Sharma et al., 2012; Yuan et al. 2012). Tradizionalmente i bivalvi sono stati suddivisi in cinque sottoclassi esistenti (Protobranchia, Pteriomorphia, Palaeoheterodonta, Heterodonta, basati principalmente sulla struttura della conchiglia e sull’anatomia). Tuttavia, una nuova classificazione è stata recentemente proposta ed adottata dal registro mondiale delle specie marine (Bieler et al., 2010). Gli autori hanno utilizzato una varietà di informazioni filogenetiche tra cui l'analisi molecolare, quella anatomica, la morfologia della conchiglia e la microstruttura, come pure la biogeografica, informazioni paleobiogeografiche e geologiche.
FINE SECONDA PARTE
AGGIUNGI UN COMMENTO [Se non desideri visualizzare il tuo commento sul tuo profilo Facebook, ricordati di togliere la spunta della ceckbox]
ARTICOLI CORRELATI
- L'EVOLUZIONE DEI MOLLUSCHI BIVALVI Parte 1
- L'EVOLUZIONE DEI MOLLUSCHI BIVALVI Parte 3
Questo articolo è protetto da Copyright © e non può essere riprodotto e diffuso tramite nessun mezzo elettronico o cartaceo senza esplicita autorizzazione scritta da parte dello staff di BiologiaMarina.eu.
Ideazione: Pierfederici Giovanni - Progetto: Pierfederici Giovanni, Castronuovo Motta Nicola.
Prima Pubblicazione 31 Lug 2006 - Le immagini dei Collaboratori detentori del Copyright © sono riproducibili solo dietro specifica autorizzazione.
Si dichiara, ai sensi della legge del 7 Marzo 2001 n. 62 che questo sito non rientra nella categoria di "Informazione periodica" in quanto viene aggiornato ad intervalli non regolari
XHTML 1.0 Transitional – CSS
