
I COCCOLITOFORI: Emiliania huxleyi (Ehux)
I Coccolitofori sono alghe unicellulari appartenenti alla divisione (o phylum) Haptophyta, caratterizzate da un esoscheletro di rivestimento sferico fatto di meravigliose placche calcaree che riflettono la luce, dette coccoliti. I Coccolitofori rappresentano esempi di fitonanoplancton di dimensioni microscopiche variabili da 5 a 35 μm, simili alle diatomee per il fatto che sono rivestiti di piccoli strati calcarei. Possiedono inoltre i flagelli come i dinoflagellati che consentono loro una limitata libertà di movimento.
La biomassa dei Coccolitofori è la più imponente nel mare. Si tratta di organismi che la maggior parte delle persone ignorano, che i trattati di storia naturale “liquidano” in poche righe e che per molto tempo sono persino sfuggiti all’osservazione: sono così piccoli che passano attraverso la stoffa dei più sottili retini da plancton. I Coccolitofori sono comparsi negli oceani alla fine del Triassico, circa 208 milioni di anni fa. Quando si sono estinti gli enormi dinosauri, la stessa sorte è toccata alla maggior parte dei Coccolitofori, ad eccezione di poche specie che erano state appena notevoli prima. Queste poche specie sopravvissute, durante i 65 milioni di anni hanno evoluto nuove forme in risposta ai cambiamenti delle condizioni ambientali. I geologi avevano osservato in certi calcari, soprattutto nelle crete, dei piccoli granuli a forma di dischi, di corone, di secchielli o di panieri senza fondo non più grandi di 1 μm che chiamarono coccoliti. Ma non sapevano che quelle particelle fossero i gusci di certi organismi. Quando scoprirono quegli organismi, li chiamarono portacoccoliti, da qui il termone Coccolitofori.
Attualmente negli oceani ne esistono circa 200 specie. I loro esoscheletri calcarei si trovano nei depositi marini spesso in gran numero, a volte costituiscono la componente principale di una particolare roccia, come quella calcarea in Gran Bretagna. I coccoliti fossili che formano le celibre scogliere bianche di Dover, in Gran Bretagna, e sono importanti fossili guida nello studio geologico degli strati di rocce sedimentarie. I Coccolitofori e in particolare la loro specie più abbondante e dominante, Emiliania huxleyi (Fig.1) sono i principali organismi calcificanti che possono formare estesi bloom in acque off-shore, dai tropici ai mari artici.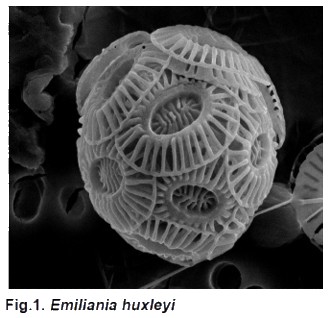
La maggior parte di questi bloom sono causati dalla specie Emiliania huxleyi, conosciuto anche come Ehux che risulta essere il più abbondante dei Coccolitofori. Attualmente, Emiliania huxleyi è una delle specie più diffuse del pianeta, presente (almeno negli strati illuminati) in una concentrazione di circa 1000 individui per litro di acqua marina. Si può rapidamente calcolare, considerando il volume delle acque della zona eufotica (dove arriva la luce), che quest'alga fitoplanctonica è presente sul pianeta con circa 6 x 1023 individui. Quando le condizioni dell’acqua sono favorevoli, Ehux ha la capacità di svilupparsi in voluminosi bloom, a volte di 100.000 km2. Durante questi bloom il numero di cellule di Ehux solitamente supera quello di tutte le altre specie, costituendo circa l’80-90% del numero totale di cellule fitoplanctoniche presenti nell’acqua. Le cellule di Ehux sono accompagnate da un gran numero di coccoliti di cui molti sono fissati alle cellule ma molti galleggiano anche nell’acqua. La presenza di questi coccoliti liberi è dovuta:
- alla sovrapproduzione di coccoliti che porta ad un numero superiore di quello che può contenere saldamente la superficie della cellula;
- alla morte cellulare dopo la quale le coccosfere vuote si disintegrano;
- alla divisione asessuale della cellula dopo la quale la coccosfera deve rompersi e rilasciare uno o entrambi gli abitanti
Le cellule di Emiliania huxleyi sono così piccole (6 μm) che al microscopio ottico appaiono solo come piccoli puntini o chicchi (in greco: cocco). Per questo motivo le cellule vive osservate in mare sono molto sparse. Con l’aiuto dei satelliti le osservazioni in mare ora sono più frequenti. Tuttavia i principi e gli interessi degli studi al satellite sono stati completamente ignorati fino al 1982. Nel 1978, al largo della Bretagna dove vengono generate altissime concentrazioni di fitoplancton in estate, è stato osservato una gamma di variazione del colore dell’oceano molto alta e quindi evidente in quella zona. Nel 1982 fu confermata la presenza di Ehux al largo della Bretagna mediante immagini del satellite CZCS. Oggi il rilevamento dei Coccolitofori è un parametro comune del SeaWiFS degli Stati Uniti e del MODIS. Inoltre, lo studio su Emiliania huxleyi e di altri Coccolitofori si è trasformato in una parte importante dell’oceanografia biologica a causa del loro ruolo nei cicli biogeochimici globali.
La struttura cellulare di Emiliania huxleyi
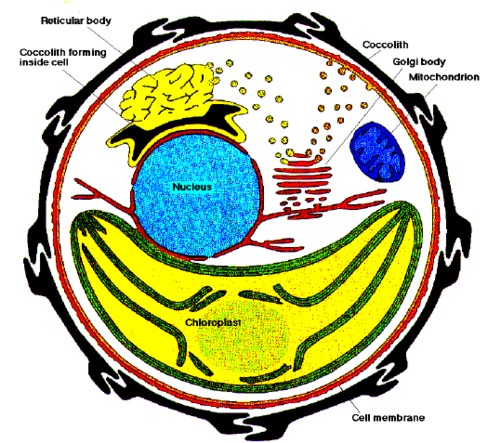
Emiliania huxleyi è unicellulare e contiene un numero di organelli interni. Il cloroplasto è quello che domina la cellula ed è il corpo dove avviene la fotosintesi. Esso contiene i pigmenti fotosintetici e funziona allo stesso modo dei cloroplasti delle piante terrestri, anche se i pigmenti differiscono per il colore giallo-bruno o dorato piuttosto che verde. Al centro della cellula, al di sopra del cloroplasto c’è il nucleo che contiene il materiale genetico e qui si ha la replica del DNA durante la divisione cellulare e la produzione di RNA. Il mitocondrio è un organello distinto, presente in tutte le cellule eucariote ed è la fonte dell’adenosina trifosfato (ATP), la principale molecola per il trasferimento di energia nei sistemi biologici. La membrana nucleare è collegata con una rete interna di membrane al reticolo endoplasmatico (ER) e attraverso questo alla membrana cellulare. Tutte queste sono formate dall’apparato del Golgi. Il cloroplasto e il mitocondrio invece hanno discrete membrane che li separano dal resto della cellula e contengono DNA separato. Ciò indica che i cloroplasti ed i mitocondri rappresentano organismi unicellulari che sono stati poi inglobati per endosimbiosi nell’ospite protista. La caratteristica peculiare della cellula di Ehux è il rivestimento dei coccoliti. I coccoliti si formano nella cellula mediante un processo altamente organizzato, detto coccolitogenesi. Durante tale processo si sviluppa un coccolito alla volta all’interno di una vescicola vicino alla membrana nucleare. Sopra la vescicola dei coccoliti c’è il corpo reticolare che controlla la formazione del coccolito. Il corpo reticolare è abbastanza separato dal corpo del Golgi ma è strettamente collegato ad esso e microvescicole possono essere osservate fra i due organelli. In altri Coccolitofori le vescicole dei coccoliti si formano all’interno del Corpo del Golgi senza la presenza di un corpo reticolare. Quando il coccolito si è formato completamente, la vescicola migra verso il bordo della cellula, le membrane della vescicola fondono con la membrana della cellula e le parti esterne della membrana sono riassorbite in modo che il coccolito è espulso fuori della cellula. Ehux, come altri Coccolitofori, possiede due flagelli lisci ed un terzo flagello, l’aptonema che è attorcigliato e ha una substruttura microtubulare abbastanza differente dai flagelli.
Il ciclo vitale
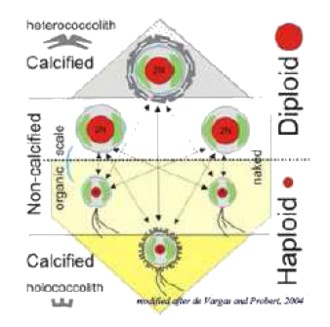
Il ciclo vitale di Emiliania huxleyi è complesso, si tratta di un ciclo aplodiplonte che consiste:
- della fase degli eterococcoliti con cellule-C diploidi, non mobili dove i coccoliti sono formati da un apparato radiale di cristalli di calcite e sono prodotti nella cellula;
- della fase degli olococcoliti con cellule-S aploidi, mobili, ricoperte con scaglie non mineralizzate.
Gli olococcoliti che sono formati da numerosi e minuscoli cristalli euedrali organizzati in allineamenti continui che si calcificano fuori alla cellula, non sono prodotti nella fase aploide. A differenza di altre specie di Coccolitofori, nel ciclo vitale di Ehux la fase alternata si presenta non-calcificata. Un terzo tipo di cellula non-calcificata, la cellula-N, è diploide e non sembra essere parte del ciclo vitale aplodiplonte, bensì un mutante. Tutti i tipi di cellule sono capaci di riproduzione asessuale indefinita, mediante scissione binaria. La fase aploide sembra essere più adattata a condizioni oligotrofiche e in alcuni casi anche mesotrofiche; la fase degli eterococcoliti invece è adattata più a condizioni eutrofiche. Inoltre, si pensa che le cellule-S abbiano una fase sessuale dove esse si comportano come gameti.
I coccoliti
Emiliania huxleyi produce coccoliti che hanno un diametro di circa 2.5 x 10-6 metri e pesano circa 1.8 x 10-12 gr. ognuno. Tipicamente c’è un multistrato di coccoliti intorno alla cellula e ogni coccolito ha circa 30 unità di cristalli. La formazione dei coccoliti avviene nella cellula e inizia con la nucleazione di un anello ellittico di semplici cristalli di calcite, un anello proto-coccolito. Questo anello proto-coccolito è situato nel punto che diventerà la base del tubo, lo sviluppo poi progredisce simultaneamente in varie direzioni.
I coccoliti di Ehux hanno la stessa struttura di base di quattro elementi formati da un unico cristallo. Per quanto concerne le funzione dei coccoliti, sono state avanzate diverse possibilità. Essi possono servire per proteggere la membrana cellulare da cambiamenti delle condizioni ambientali, dall’eccesso di luce ultravioletta o per proteggere da eventuali traumi fisici. Inoltre, i coccoliti possono scoraggiare i predatori in quanto le placche indigeribili corazzano le cellule e riducono il loro valore nutrizionale, forse rendendo i coccolitofori un po’ meno appetibili. 
I coccoliti intrappolano l’acqua, creando una zona respingente in cui la cellula ha un più alto controllo sulla chimica dell’acqua. In Emiliania huxleyi la formazione dei coccoliti potrebbe aiutare anche la fotosintesi. La reazione chimica usata per formare carbonato di calcio rilascia anche anidride carbonica nella cellula dove può essere consumato immediatamente. I coccoliti possono essere utilizzati anche per ridurre la velocità di affondamento in caso di acque calme; nel caso di acque turbolenti, invece, i coccoliti possono aumentare la densità della cellula in modo da favorire l’affondamento e quindi l’accesso di nutrienti. I coccoliti possono regolare anche la luce che entra nella cellula.
Sopra, a sinistra, parte interna di un coccolito di Ehux, a destra la parte superiore.
Che cosa fanno all'ambiente?
I Coccolitofori normalmente non risultano essere nocivi per la vita degli altri organismi nell'oceano. Nelle zone povere di nutrienti dove altro fitoplancton è limitato, i Coccolitofori sono una fonte di nutrizione per molti piccoli pesci e zooplancton che mangiano altri organismi fitoplanctonici. Quando i Coccolitofori sono presenti in gran numero possono bloccare le branchie dei pesci. I Coccolitofori costruiscono come detto i loro coccoliti utilizzando carbonio, calcio e tre molecole di ossigeno (CaCO3). Così ogni volta che viene formato un coccolito, un atomo di carbonio in meno vaga liberamente in atmosfera. Trecento venti libbre di carbonio entrano in ogni tonnellata di coccoliti prodotti. Alla morte tutto questo materiale affonda sul fondo dell'oceano e forma nuovo sedimento. L'effetto dei Coccolitofori sull'ambiente è piuttosto complesso. Questo effetto riguarda ancora la formazione dei loro coccoliti e la reazione chimica che si ha durante tale processo. La reazione chimica che porta alla formazione dei coccoliti inoltre genera una molecola di anidride carbonica, un gas serra molto potente a partire dall'ossigeno e dal carbonio già presente nell'oceano. Mentre gran parte del gas entra nei coccoliti, un’altra parte va nell'atmosfera ed immediatamente si trasforma contribuendo al problema dei gas serra. Gli scienziati sostengono che a breve termine i gas serra faranno diventare gli strati superiori dell'oceano più temperati e stagnanti. Ciò aumenterebbe il numero di coccoliti nel mondo e quindi ci sarebbe più produzione di gas serra. L'accumulo di anidride carbonica nell'atmosfera, oltre a causare il riscaldamento della superficie terrestre è responsabile di un'altra minaccia all'ambiente globale: sta provocando l'acidificazione degli oceani, cosa che rischia di impedire la riproduzione dei coralli e di altre specie che aiutano i mari ad assorbire anidride carbonica. Tuttavia è stato dimostrato che questi microrganismi non possono vivere e riprodursi in un mare più acido. Continuando quindi a immettere gas serra nell'atmosfera ai livelli attuali, gli esperti hanno previsto che il pH dell'acqua marina, normalmente basica, diminuirà sensibilmente e ciò causerà la scomparsa dei Coccolitofori, bloccando la loro funzione di ricettori di carbonio dall'atmosfera e compromettendo irrimediabilmente la capacità filtrante dei mari di altre specie. La situazione quindi sta evolvendo verso un pericoloso circolo vizioso: lo scarico di anidirde carbonica nell'aria diminuisce la capacità di assorbimento dei mari che, a sua volta, aumenta la quantità complessiva di anidride carbonica presente nell'atmosfera, contribuendo ad accelerare il riscaldamento della terra. I Coccolitofori inoltre hanno influenza sul clima globale in quanto aumentano l'albedo degli oceani. L'albedo è la frazione di luce solare che un oggetto riflette (più alti valori di albedo indicano più luce riflessa). I bloom dei Coccolitofori riflettono quasi tutta la luce visibile che li colpisce. Poiché la maggior parte di questa luce viene riflessa, solo una piccola parte viene assorbita dall'oceano e immagazzinata come calore.
A differenza di altri organismi fitoplanctonici che vivono sulla superficie dell’oceano dove c’è acqua più fredda e abbondanza di sostanze nutritive (upwelling), i Coccolitofori preferiscono vivere negli strati superficiali in acque poco profonde, povere di nutrienti con temperature più calde. Inoltre, i Coccolitofori non hanno bisogno di un afflusso costante di nutrienti per vivere. Prosperano spesso nelle zone dove i loro competitori sono affamati e una volta che si sviluppano in una regione, diventano dominanti fino a costituire circa il 90% del fitoplancton di quella zona. I Coccolitofori si sviluppano principalmente nelle regioni subpolari, alle latitudini temperate. Altri posti in cui possono presentarsi bloom di Coccolitofori sono il litorale nordico dell'Australia e le acque che circondano l'Islanda. Negli ultimi anni i grandi bloom di Coccolitofori sono stati rilevati anche nel mare di Bering. Ciò ha sorpreso molti scienziati poiché il mare di Bering è normalmente un corpo d’acqua ricco di nutrienti. Questi microfitoplancton costituiscono un componente chiave nel ciclo dello zolfo oceano-atmosfera attraverso il rilascio di dimetilsolfuro (DMS), noto come la principale risorsa naturale di zolfo atmosferico e considerato uno dei regolatori chiave del clima quotidiano. Il DMS è repulsivo per i pesci che possono allora cambiare la rotta di migrazione.
Altri due prodotti dei bloom di Coccolitofori sono l’esportazione del carbonato di calcio nelle profondità dell’oceano e il rilascio di CO2 in superficie. Questo flusso biogenico di carbonato che affonda nell’oceano costituisce circa il 60% del flusso totale di carbonio che va nei sedimenti del mondo, spesso paragonabile al carbonio organico prodotto e accumulato soprattutto nelle zone oligotrofiche e mesotrofiche.
Ecologia di Emiliania huxleyi
I bloom di Ehux si presentano in acque molto stratificate dove la profondità dello strato superficiale mescolato è generalmente ~10-20 m e quasi sempre <= 30m. Ciò suggerisce che l’elevata intensità della luce può causare la formazione dei bloom a metà estate quando le radiazioni di superficie sono alte. Un’ulteriore prova per questa ipotesi viene dagli esperimenti del mesocosmo condotti da vari anni nei fiordi Norvegesi. Un’analisi dei vari parametri ambientali che possono influenzare la formazione e distribuzione della popolazione di Ehux, ha indicato che l’elevata irradianza di superficie è il parametro più influente. È stato visto inoltre che l’esposizione di Ehux ad un’alta intensità luminosa non provoca fotoinibizione. I risultati degli esperimenti del mesocosmo nei fiordi Norvegesi hanno suggerito che Ehux può presentare un vantaggio competitivo rispetto ad altri organismi fitoplanctonici quando il fosfato inorganico è limitato mentre il nitrato è più abbondante. Con questi esperimenti si è anche concluso quindi che lo sviluppo di Ehux potrebbe essere spiegato meglio assumendo che questa alga è più capace nell’utilizzo di fosfati organici disciolti rispetto agli altri organismi fitoplanctonici. È stato dimostrato che c’è un’attività dell’enzima fosfatasi alcalina in Ehux molto più grande rispetto alla maggior parte degli altri organismi fitoplanctonici e che questo enzima viene usato per assimilare fosfati organici disciolti. Uno studio effettuato nel Nord-Est Atlantico ha dimostrato che si ha un più alto numero di Ehux quando c’è basso fosfato ed elevata luminosità. La disponibilità di CO2 nell’acqua gioca un ruolo nella determinazione delle latitudine e dei periodi dell’anno in cui Ehux può diventare dominante. I Coccolitofori, come Ehux, possiedono un vantaggio competitivo a basse concentrazioni di CO2 a causa del processo di calcificazione che sintetizza CO2 nella cellula a partire da HCO3-.
È stato dimostrato anche che una bassa concentrazione di silicati può essere una possibile spiegazione della distribuzione biogeografia di Ehux. Alcune altre ipotesi sono: l’influenza causata da “effetti di semina” (è richiesto il trasporto di cellule per dare inizio alla popolazione prima che si possa avere un bloom); un vantaggio a bassa irradianza e basso nitrato/ammonio, e un’alta tolleranza a basse concentrazioni di ferro. Inoltre, Ehux ha bisogno di tiamina (vitamina B1) la quale non è presente in acqua se non c’è attività biologica. Pertanto i bloom di Ehux sono possibili solo se bloom di altre specie fitoplanctoniche, quale quelli delle diatomee, li precedono condizionando l’acqua. È stato visto che nel Mar del Nord i bloom di Ehux, infatti, seguono quelli delle diatomee in acque che sono state impoverite di nutrienti inorganici e sono diventate più stabili in termini di mescolamento verticale (per esempio in seguito alla diminuzione di upwelling o alla formazione del termoclino stagionale). È stato notato anche che lungo la costa sud-ovest della Norvegia i bloom di Emiliania huxleyi che colorano il mare di verde latteo seguono annualmente i bloom di diatomee in primavera.
Impatti biogeochimici
I bloom di Emiliania huxleyi possono essere molto importanti in termini di temperatura globale del pianeta. Quando questi bloom compaiono sopra le ampie estensioni dell’oceano, essi hanno innumerevoli effetti sull’acqua e sull’atmosfera. Sebbene ogni cellula è invisibile, possono esserci un migliaio di miliardi (1021) di cellule in un ampio bloom e la popolazione nell’insieme ha un enorme impatto. Il lavoro su questo impatto è difficile prima di tutto per valutare se l’effetto dei bloom è quello di esacerbare o migliorare il riscaldamento globale. I bloom di Ehux sono processati attraverso la catena alimentare marina, con i virus, i batteri e lo zooplancton che contribuiscono tutti al trasferimento e alla decomposizione dei bloom. Alcuni detriti dei bloom sopravvivono affondando sul fondo dell’oceano e rilasciano prodotti chimici lungo la colonna d’acqua. Durante la loro vita e quando muoiono (Fig.2), le cellule del fitoplancton disperdono sostanze chimiche nell’acqua.
Un bloom può essere immaginato sia come una fabbrica compatta di sostanze chimiche che rimuove dall’acqua anidride carbonica, nitrati, fosfati e altri nutrienti disciolti, e allo stesso tempo immette nell’acqua altre sostanze chimiche come ossigeno, ammoniaca, dimetilsolfuro (DMS), che viene fornito direttamente all’atmosfera, e altri composti organici disciolti. Allo stesso tempo la fabbrica di sostanze chimiche pompa grandi volumi di materia organica e carbonato di calcio (CaCO3) lungo la profondità dell’oceano e sul fondo. Alcuni di questi carbonati di calcio finiscono come gesso o calcare nelle rocce sedimentarie marine, forse per passare attraverso la crosta terrestre e riapparire milioni di anni dopo come montagne, colline e scogliere. Uno studio globale effettuato mediante il satellite ha rilevato un’area annuale di bloom pari a 1,4 milioni di Km2. Questa è l’area totale che ogni anno è stata classificata come bloom di Coccolitofori, cioè quella che è oscurata dalle nubi e soddisfa le soglie imposte in termini di luminosità, dimensione dell’area del bloom ed altri parametri. Inoltre c’è una produttività di Coccolitofori in aree dove non ci sono bloom, per esempio nell’area oligotrofica degli oceani tropicali, dove c’è un continuo ambiente di turnover dei Coccolitofori anche se la loro abbondanza non raggiunge mai le proporzioni dei bloom.
Siccome i flussi di calcite non sono determinati soltanto da quelli di Ehux, sono stati considerati gli impatti biogeochimici della produttività globale di calcite nell’oceano, cioè la produttività di Ehux ed altri Coccolitofori e anche di altri organismi. Sebbene Ehux è frequentemente il coccolitoforo più abbondante in campioni d’acqua di mare, a causa delle sue piccole dimensioni non sempre domina l’afflusso di calcite. Dalla produttività stimata, dai volumi di sedimento valutato e dai modelli geochimici, si è potuto stimare quanto carbonato di calcio (calcite) viene prodotto annualmente dai coccolitofori, foraminiferi e dai banchi di coralli (i principali produttori di calcite negli oceani). È stato stimato che la produzione totale di calcite è in media di un miliardo di tonnellate all' anno. La rispettiva contribuzione dei coccolitofori, dei foraminiferi e di altri organismi alla produzione totale di calcite rimane relativamente poco conosciuta.

Fig. 2. Coccoliti ammassati dopo un bloom di Emiliana huxleyi
Una significativa percentuale della calcite prodotta (in qualche zona tra lo 0 e l’80%) probabilmente si dissolve prima dell’incorporazione nei sedimenti. Ciò fa nascere la domanda: quanta calcite viene deposta? Questo dato è più facile da ottenere, dalla media della quantità di calcite in diversi centri di sedimenti marini e dividendo con il livello di accumulazione del materiale nei centri. È stato stimato un livello di accumulazione di carbonato di calcio sedimentario pari a 0.46 Gt di calcite in un anno. Molta della calcite disciolta (la differenza tra produzione e sepoltura) probabilmente è presente sulla superficie delle acque, ma una buona quantità è presente anche sui grandi fondali. Dal confronto tra le quantità di calcite in diversi centri di sedimenti marini, è risultato che alcune parti del fondale marino non sono affatto raggiunte da calcite, soprattutto il fondale marino sotto le acque più profonde. Questo a causa di ciò che è conosciuto come calcite lisoclina. Al di sotto di una certa profondità (circa 4-5 km) le pressioni alte causano un cambiamento nella chimica dell’acqua tale che la calcite viene forzata in soluzione. L’acqua improvvisamente diventa molto più corrosiva per la calcite e tutti i frammenti di calcite che piovono verso il fondo del mare si dissolvono rapidamente al di sotto della profondità critica del lisoclino.
Emiliania huxleyi gioca un ruolo importante sul clima della Terra sia attraverso la formazione dei coccoliti cioè delle placche di carbonato le quali rendono l’oceano opalescente, sia attraverso l’emissione di gas. Essa altera quindi il clima, aumentando la copertura delle nubi mediante particelle di aerosol solfati prodotte dal dimetisolfuro emesso per mantenere il bilancio ionico contro il sodio. Ci sono diversi modi con cui i Coccolitofori influenzano la temperatura locale e globale:
1) Albedo dell’oceano I coccoliti agiscono un po’ come piccoli specchi a causa delle speciali proprietà ottiche della calcite. Questo causa un tipico bloom (che contiene 100 mg/ m3 di carbonio della calcite) che aumenta l’albedo dell’oceano dal ~7.5% al ~9.7% (Fig.3). Un altro tipo di modello ottico (più accurato per questo scopo) calcola un aumento dal ~6.2% al ~9.7%. Se si assume che ogni bloom persiste per circa un mese, allora una copertura annuale di 1.4 x 106 Km2 aumenterebbe il globale albedo planetario annuale medio dello (9.7 - 6.2) x (1/12) x (1.4/510) = 0.001% dove 510 è l’area della Terra in 106 Km2. Questo è il limite più basso sull’impatto totale in quanto la concentrazione di coccoliti del sub-bloom che diffondono la luce avrà un effetto come bloom oscurato da nubi su aree molto più ampie (impatto massimo di albedo stimato=0.21%), le quali non sono incluse nell’area annuale totale di bloom in quanto potrebbero non essere viste dal satellite. Un cambiamento di albedo dello 0.001% corrisponde ad una riduzione di 0.002 W/m2 dell’energia solare in entrata, mentre un cambiamento di albedo dello 0.21% causa una riduzione di 0.35 W/m2.
Questi due numeri possono essere paragonati all’influenza dell’aggiunta antropogenica di anidride carbonica dal 1700 che è stata stimata essere circa 2.5 W/m2. I coccoliti che diffondono la luce sono perciò solo un fattore di importanza secondaria nel bilancio radioattivo della Terra.
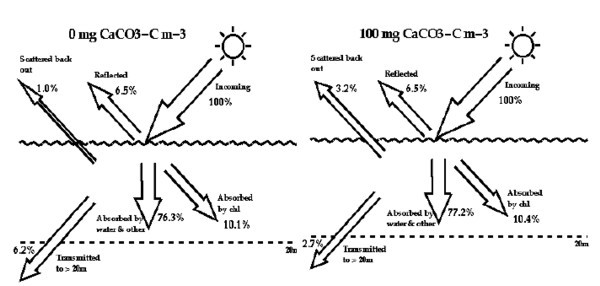
Fig.3. Bilancio di fotoni per acqua senza coccoliti (a sinistra) e per acqua con una concentrazione di 100 mg/m3 di carbonio della calcite nella forma di coccoliti.
2) Ritenzione di calore dell’oceano La diffusione della luce causata dai coccoliti apporta più calore e luce che generalmente vengono spinti di nuovo nell’atmosfera. Inoltre ciò fa sì che gran parte del calore restante viene bloccato in prossimità della superficie dell’oceano e soltanto una piccolissima frazione del calore totale riesce a penetrare più in profondità. Siccome lo scambio di calore con l’atmosfera avviene in superficie, tutti e tre gli effetti appena descritti contribuiscono a voler dire che i bloom dei Coccolitofori possono far raffreddare drasticamente tutta la colonna d’acqua durante un lungo periodo, anche se ciò potrebbe essere mascherato inizialmente da un riscaldamento della superficie dell’oceano (pochi metri). Attualmente si sta studiando l’importanza di questo effetto sia a livello regionale che globale ma non è stata ancora stabilita.
3) Albedo delle nubi Il fitoplancton produce un composto dello zolfo, il dimetilsolfuro DMS (CH3-S-CH3), con i Coccolitofori e altre specie che ne producono 100 volte di più rispetto alle diatomee. Il DMS è una delle più importanti tracce gassose emesse dagli oceani nell’atmosfera. Il DMS viene prodotto dalla scomposizione di una molecola detta dimetilsolfoniopropionato (DMSP) che si trova nelle cellule di alcune specie di fitoplancton. Si pensava che il fitoplancton produceva DMSP per la sua sopravvivenza alle condizioni di salinità negli oceani, ma è stato visto che gli organismi fitoplanctonici possono usare DMSP anche per eliminare i rifiuti dannosi, per permettersi di vivere in acque molto fredde e forse persino per evitare che altri animali li mangiano. I Coccolitofori sono importanti produttori di DMSP. Quando le cellule del fitoplancton vengono danneggiate, per esempio da turbolenze, dal grazing dello zooplancton o da lisi virali e quindi muoiono, il DMSP viene rilasciato in mare e si scompone a formare il DMS. Dopo la morte delle cellule e a successive trasformazioni biologiche e chimiche nell’oceano e nell’atmosfera, gran parte del DMS rimane in acqua ed è consumato dai batteri o è convertito in altre specie chimiche, mentre una parte del DMS fuoriesce e diffonde dal mare all’atmosfera come precursore dei nuclei di condensazione delle nubi (CCN). Questi CCN aiutano a stimolare la formazione delle nubi, soprattutto nelle aree dove altre risorse di CCN sono in piccola scorta e la quantità di CCN limita il possibile sviluppo delle nubi. Una volta trasferito all’atmosfera il DMS gassoso viene foto-ossidato a aerosol solfati troposferici i quali agiscono come nuclei di condensazione delle nubi (CCN) che attraggono molecole d’acqua. Il vapore d’acqua condensa su queste particelle di CCN e formano le goccioline d’acqua che compongono le nubi. Nell’atmosfera l’acqua è presente sotto forma di vapore che condensando determina la formazione delle gocce. Il DMS gassoso viene foto-ossidato formando anidride solforosa (SO2) la quale reagisce con microscopiche particelle metalliche che catalizzano, con il contributo delle radiazioni solari, la reazione che porta alla formazione dell’anidride solforica (SO3). L’anidride solforosa e l’anidride solforica possono ossidarsi in combinazione con l’acqua formando acido solforico (H2SO4) che provoca fenomeni di acidificazione (piogge acide).
In questo modo la produttività dei Coccolitofori aiuta ad aumentare la capacità di riflessione (albedo) del pianeta favorendo la formazione di più nubi. Le nubi intaccano il bilancio radioattivo della Terra e perciò influenzano molto la sua temperatura ed il clima. Il DMS rappresenta il 95% del naturale flusso marino di gas di zolfo all’atmosfera e gli scienziati stimano che il flusso di DMS marino fornisce circa il 50% della risorsa biogenica globale di zolfo all’atmosfera. Il DMS, quindi, è il composto di zolfo biogenico dominante nell’atmosfera marina ed è essenziale per il ciclo globale dello zolfo. Un processo chiave nel ciclo dello zolfo è il trasferimento di composti di zolfo volatili dal mare alla terra attraverso l’atmosfera. C’è una relazione tra DMS, aerosol solfati e nuclei di condensazione delle nubi. Siccome gli aerosol solfati funzionano come nuclei di condensazione delle nubi, il DMS ha un effetto significativo sul clima della Terra. Il DMS prodotto dal plancton e il suo trasferimento all’atmosfera è considerato uno dei meccanismi mediante il quale il biota può regolare il clima. Il rilascio di DMS, quindi, può agire come governatore del clima globale. Siccome il DMS è il prodotto della scomposizione di un composto chimico prodotto biologicamente, le emissioni di DMS si presentano in primavera, estate ed autunno quando il fitoplancton si sviluppa. Le emissioni provengono sia dalle acque costiere che dall’oceano aperto ma sono maggiori nelle regioni dove il fitoplancton si sviluppa particolarmente. Il DMS viene emesso a nord-est dell’Oceano Atlantico e dal Mare di Bering a nord dell’Oceano Pacifico (Fig.4) in quanto in queste aree ci sono bloom regolari di coccolitofori. Inoltre il DMS entra nell’atmosfera anche dalle acque costiere intorno all’Europa. La quantità di zolfo emessa dalle fonti naturali è quasi uguale a quella prodotta durante la combustione di combustibili fossili operata dagli esseri umani. Pertanto il fitoplancton è importante al nostro clima quanto le enormi centrali elettriche che generano la nostra energia. Nelle zone lontane dall’attività umana, la maggior parte dello zolfo presente nell’atmosfera deriva dal DMS emesso dal mare. È stato stimato che gli oceani forniscono all’atmosfera da 20 a 50 tonnellate di zolfo ogni anno.

Fig.4. Bloom visibili dai satelliti nello spazio. Il colore verdastro è un bloom del coccolitoforo nel Mare di Bering visto dal satellite SeaWiFS della NASA.
4) Il gas serra CO2 Emiliania huxleyi, come gli altri Coccolitofori, sono importanti produttori primari nell’oceano alle latitudini temperate ma sono anche tra i maggiori trasportatori di carbonio inorganico dalla superficie al fondo del mare. I Coccolitofori sono coinvolti nello scambio di anidride carbonica tra l’atmosfera e l’oceano mediante la fotosintesi e i loro gusci di rivestimento in carbonato di calcio aumentano i livelli di carbonio organico prodotto da altri produttori primari nella zona fotica pertanto hanno un ruolo rilevante nel ciclo biogeochimico globale del carbonio.
Lo sviluppo del fitoplancton trasforma la CO2 in materia organica e riduce la CO2 atmosferica. Tuttavia i Coccolitofori sono unici in quanto essi prendono anche il bicarbonato (HCO3-) con cui formare il carbonato di calcio dei loro coccoliti. La reazione chimica per la formazione dei coccoliti è:
Ca2+ + 2HCO3- ---------> CaCO3 + H2O + CO2
Ci sono tre forme di carbonio inorganico disciolto nel mare: gas disciolto, CO2; ione bicarbonato, HCO3- e ione carbonato, CO32-; e il carbonio può cambiare facilmente dallo stato di una di queste forme disciolte in altri stati. La quantità di carbonio totale presente in ogni forma è determinata dall’alcalinità e dalla temperatura dell’acqua. Quando il sistema del carbonio in mare è perturbato dalle cellule dei coccolitofori che rimuovono HCO3- per formare i coccoliti, questo causa un riarrangiamento più o meno istantaneamente della quantità di carbonio presente in ogni forma disciolta. La rimozione di 2 molecole di HCO3- e l’aggiunta di una molecola di CO2 cambia l’alcalinità e ciò indirettamente causa che la maggior parte del carbonio disciolto è presente nella forma CO2. Sebbene il carbonio disciolto totale è ovviamente ridotto mediante la rimozione di carbonio disciolto (ioni bicarbonato) nel carbonato di calcio solido, tuttavia l’effetto totale, paradossalmente, è la produzione di più CO2 disciolto in acqua. In questo modo i bloom dei coccolitofori tendono ad esacerbare il riscaldamento globale (causando l’aumento di CO2 atmosferica), piuttosto che migliorarlo come avviene quando la CO2 disciolta entra nella nuova biomassa organica. Tuttavia un lavoro recente ha mostrato che le proprietà supplementari dei coccoliti possono rendere la situazione ancora più complessa. La calcite dei coccoliti è piuttosto densa (2.7 Kg per litro rispetto alla densità del mare di 1.024 kg per litro) e la presenza dei coccoliti nelle palline fecali dello zooplancton e nella “neve marina” (la chitina, un polisaccaride che viene degradato dall’azione batterica in mare dopo che sugli stessi batteri arrivano segnali chimici) (le due principali forme in cui la materia biogenica affonda lungo la profondità dell’oceano) li induce ad affondare più velocemente. Con l’affondamento lento la materia organica può aderire anche alla superficie dei coccoliti che galleggiano sulla superficie delle acque. Se la materia organica affonda più velocemente allora c’è meno tempo per essere attaccata dai batteri e in questo modo la maggior parte del carbonio bloccato può fuoriuscire dalle acque superficiali, esaurendo la CO2 di superficie. Uno studio recente ha dimostrato che questo co-trasporto della materia organica con i coccoliti controbilancia l’aumento di CO2, che diversamente sarebbe provocato, e fa agire i bloom dei Coccolitofori per opporre il riscaldamento globale piuttosto che per intensificarlo.
La biogeografia di Emiliania huxleyi
I bloom di Ehux sono stati osservati nelle seguenti zone:
- Oceano Atlantico del Nord
- Golfo del Maine
- Penisola della Nuova Scozia (Canada)
- Mar del Nord (come nello Skagerrak)
- Fiordi Norvegesi
- Arcipelago delle Bermuda
- Baia di Biscaglia (Oceano Atlantico nord-orientale)
- Mare di Bering (Oceano Pacifico)
- Mare di Weddell (Antartide)
- Mar Nero
- Vicino all’Isola di Scilly (a sud della Gran Bretagna)
- Sud del Bengala (Oceano Indiano)
Tutti i bloom di Ehux conosciuti presentano una caratteristica idrografica comune: essi si presentano tutti in acque altamente stratificate dove la profondità in genere è di ~10 - 20 m. ed è quasi sempre <= 30m. Una seconda fonte molto importante per le informazioni sulla biogeografia di Ehux viene dalle immagini proveniente dal satellite. Grazie ai satelliti si conosce molto di più circa la distribuzione dei bloom di Ehux che di altre specie.
La riflessione della luce mediante i coccoliti significa che le immagini dei satelliti possono dare un'immagine ad alta definizione istantanea della diffusione totale in modo dimostrabile del bloom di una singola specie, che non è possibile per altre specie. Uno studio ha analizzato un gruppo di dati globali presi da immagini del satellite CZCS durante gli anni 1979 - 1985. E’ stata costruita una procedura per rilevare i bloom dei Coccolitofori (i quali risultarono quasi tutti di Ehux) e poi i dati ottenuti in più anni sono stati applicati sul globale per avere una valutazione della distribuzione globale delle specie.

Bloom di Ehux: A) Mar Celtico; B) a ovest dell’Alaska; C) Isola di Terranova Atlantico occidentale
Distribuzione globale dei bloom di Emiliania huxleyi
I bloom dei coccolitofori Emiliania huxleyi localmente agiscono come un’importante risorsa di dimetilsolfuro (DMS) e di carbonato di calcio ed alterano le proprietà ottiche dello strato superficiale. Questi bloom spesso ricoprono vaste aree e possono essere identificati dalle immagini provenienti dal satellite a causa della grande quantità di luce riflessa dalla colonna d’acqua. La loro presenza dà all’oceano un’apparenza bianco latte/turchese. La capacità di rilevare i bloom di Emiliania huxleyi nelle immagini provenienti dal satellite, oltre che fornire la conoscenza biogeografica della specie nel tempo e nello spazio, fornisce un metodo per valutare la loro importanza biogeochimica sulla base di scale globali. Il CZCS (Coastal Zone Color Scanner) è stato usato per mappare la distribuzione dei bloom di Ehux e per valutare la grandezza e la periodicità della loro produzione di CaCO3 e di DMS negli oceani. I pixel delle immagini composite di 5 giorni dell’intera missione di CZCS sono stati classificati in classi di bloom e classi di non-bloom basati sulla loro radiosità media lasciata sull’acqua usando uno schema sorvegliato e multispettrale. È stata messa a punto una procedura che mette a confronto le caratteristiche spettrali note dei bloom di Ehux e quelle delle condizioni di nonbloom. Le caratteristiche spettrali dei bloom di Ehux, acqua blu “trasparente”, sedimento caricato dell’acqua, “gesso in polvere” (fanghi sospesi della calce), e nebbia atmosferica sono state estratte dalle immagini del CZCS. Un insieme di dati indipendenti inoltre sono stati usati per stabilire che la procedura fosse efficace per distinguere i bloom dei coccolitofori da altre condizioni dell’acqua, con l’eccezione dei “gessi in polvere”, alla risoluzione spaziale dell’immagine globale. Le immagini classificate mediante lo schema sono state raccolte nei mensili, negli annuali e nelle missioni di climatologia delle zone di bloom e non-bloom.
Caratteristiche spettrali simili a quelle dei bloom di Emiliania huxleyi sono state trovate molto estese alle latitudini subpolari, soprattutto nelle acque superficiali del Nord Atlantico, del Nord Pacifico e della piattaforma e scarpata continentale dell’ Argentina. I bloom classificati ricoprono una media annuale di 1.4 x 106 km2, con le latitudini subpolari che stimano il 71% di questa area. I bloom individuati a queste alte latitudini rappresentano la presenza di bloom di Ehux in quanto lo schema di classificazione è risultato efficiente in queste regioni e le loro posizioni sono sostenute da indagini biogeografiche precedenti. Numerosi bloom classificati, spesso abbastanza estesi, sono stati identificati anche in mari di confine alle basse latitudini, sebbene le condizioni responsabili per questo segnale sono ambigue. A secondo della stagione, i bloom classificati nelle regioni oceaniche subpolari presentano la loro estensione spaziale più grande dall’estate a inizio autunno, mentre quelli alle latitudini più basse hanno il loro picco da metà inverno a inizio primavera. La quantità di calcite-carbonio e di DMS-zolfo prodotti dai bloom di Ehux classificati è stata valutata usando l’area di distribuzione media annuale dei bloom e i valori rappresentativi dello strato coinvolto, le concentrazioni medie delle cellule trovate nei bloom, la concentrazione di DMSP per cellula e la massa di calcite per coccolito. I bloom rilevati alle alte latitudini producono in media 0.4 – 1.3 x 106 tonnellate metriche annue di CaCO3-C e 10.000 tonnellate annue di DMS-S.
Queste valutazioni suggeriscono che i bloom rilevati dal satellite giocano solo un ruolo secondario nella produzione annuale di calcite e DMS su una scala globale. Anche se nessun satellite sensore di colore dell’oceano ha operato dopo la missione del CZCS nel mese di giugno del 1986, missioni successive come quelle col satellite SeaWiFS hanno permesso di controllare la distribuzione dei bloom di Ehux. Queste missioni dedicate al colore dell’oceano, insieme alle tecniche di valutazione dei coccoliti e delle concentrazioni delle cellule nei bloom dei coccolitofori dalle immagini provenienti dal satellite, miglioreranno la capacità di valutare l’effetto dei bloom dei coccolitofori sul ciclo del carbonio e dello zolfo in futuro. La procedura è stata aggiornata per comprendere le differenze tra le immagini del CZCS e quelle del SeaWiFS. Tuttavia vi sono delle limitazioni legate all’esistenza di condizioni che possono imitare la caratteristica spettrale dei bloom di Ehux e di conseguenza queste condizioni possono essere classificate in modo sbagliato come bloom di Ehux.
Impatti ottici di Emiliania huxleyi
Per la maggior parte dei tipi di fitoplancton sono i cloroplasti (la parte fotosintetizzante della cellula) che causano la maggiore differenza alla luce, assorbono i fotoni ed intrappolano l’energia chimica utilizzata dalla cellula. Per le specie di coccolitofori, i coccoliti così come i cloroplasti sono otticamente importanti. I coccoliti non assorbono i fotoni i quali terminano il loro percorso attraverso l’acqua, bensì si comportano come piccoli specchi che riflettono o diffondono i fotoni che causano deviazioni nella loro direzione di percorso. Ecco perché i bloom dei Coccolitofori cambiano la luminosità dell’acqua e perché i bloom possono essere visti dagli shuttle spaziali e dai satelliti. L’effetto illuminante liberato dalla massa di coccoliti è stato visto in modo sorprendente anche seguendo un esperimento artificiale (chiamato “Chalk-Ex”) in cui le rocce di gesso (i coccoliti fossilizzati!) sono state macinate prima di essere rilasciate sulla superficie dell’oceano (Fig.5) Nell’esperimento “Chalk-Ex” sono stati versati 13 tonnellate di gesso polverizzato in una zona di 2 km2 nel Golfo del Maine (Novembre 2001). 
Il gesso in polvere è stato ottenuto macinando rocce dell’età Cretacea fatte da gesso di coccoliti, spedite soprattutto dal Regno Unito. La dimensione media delle particelle era di circa 2 μm, lo stesso formato dei coccoliti di Ehux! A confronto dei tipici bloom naturali di Ehux consistono di centinaia di migliaia di tonnellate di gesso, sparsi per una più ampia zona. Anche se ogni coccolito è invisibilmente piccolo, quando sono presenti in grande numero nell’acqua causano grandi cambiamenti nel senso che la luce è trasmessa o riflessa dall’acqua. La figura 6 mostra l’effetto che differenti componenti hanno sulle proprietà ottiche dell’acqua, per esempio sulle probabilità di assorbimento (termine) e di dispersione (riflessione) dei fotoni. È possibile vedere come un intenso bloom di Ehux (100 mg di CaCO3-C per m3) causa un grande aumento nella riflessione a tutte le lunghezze d’onda (400-700nm) relative al fitoplancton, ma nessun cambiamento all’assorbimento. 400-700 nm sono anche parte dello spettro del visibile, con la luce rossa a ~650 nm, la luce del verde a ~550 nm e la luce del blu a ~450 nm.
Fig.5. Il colore pallido dell’acqua ai lati della scia della nave
Le misteriose “Acque Bianche”
Emiliania huxleyi si trova soprattutto in acque fredde dove i nutrienti sono abbondanti come nei fiordi Norvegesi. In condizioni ottimali le popolazioni di Ehux esplodono e formano enormi bloom che possono arrivare a ricoprire più di 100.000 km2 della superficie oceanica. I coccoliti liberi rilasciati dalle cellule viventi sono responsabili dell’insolito colore dell’acqua: essi diffondono la luce solare causando un cambiamento nel colore delle acque che diventano un turchese bianco-latteo (Fig.7), quasi magico in quanto incandescente e vengono chiamate “Acque Bianche”. L’acqua assume le stesse caratteristiche ottiche del latte scremato. I bloom si possono estendere da 0 a 30 metri, al di sotto di questa profondità l’acqua può essere più chiara ma molto cupo.
Il colore latteo dell’acqua è creato dall’elevata capacità di riflessione dei microgranuli di zolfo precipitato che derivano dall’ossidazione degli ioni solfato vicino alla superficie ossigenata del mare. Nelle acque profonde l’ossigeno viene consumato attraverso la respirazione batterica. In queste condizioni anossiche i batteri anaerobici utilizzano ioni solfato per ossidare la materia organica. Nel processo gli atomi di zolfo degli ioni solfato sono ridotti a solfuro che a sua volta si combina con l’idrogeno a formare idrogeno solforato. Col tempo l’idrogeno solforato si accumula nei sedimenti sul fondo del mare e un gran volume di questo gas viene rilasciato dal fondo e risale in superficie. In superficie dove è presente l’ossigeno, l’idrogeno solforato viene ossidato a zolfo elementare con conseguente formazione di una massa di particelle. La diffusione delle radiazioni solari mediante queste particelle causa un aumento delle irradiazioni che escono dall’acqua e rende visibili dallo spazio le distese dei bloom di Ehux da turchese a bianco latteo.
Oggi gli enormi bloom sono facilmente rilevabili dai satelliti a causa della quantità di luce che essi riflettono nello spazio. I bloom di Emiliania huxleyi possono contenere milioni di cellule per litro e generare decine di migliaia di tonnellate metriche di carbonato di calcio nello strato ricoperto da un solo bloom.
Il futuro di Emiliania huxleyi
Gli enormi importi di CO2 nell’atmosfera hanno influenza sul clima globale. Ma non sono gli effetti climatici ad arrecare problemi ad Ehux bensì la CO2 extra che va nell’atmosfera la quale passa alla superficie del mare ed entra così nell’oceano, aumentando il livello di carbonio disciolto. La CO2 extra disciolta ha l’effetto di acidificare l’oceano, abbassando il pH e ciò potrebbe costituire un grave problema per Ehux e anche per altri organismi calcificanti.
Gli organismi calcificanti quindi sono nei pasticci!
La possibilità con cui Ehux può formare le strutture di carbonato di calcio (CaCO3), cioè i coccoliti, è legata alla chimica dell’oceano e più precisamente a come i substrati disciolti salteranno dalla soluzione al CaCO3 solido. I substrati per il carbonato di calcio sono lo ione calcio (Ca2+) e lo ione bicarbonato (HCO3-). La distribuzione delle tre forme di carbonio inorganico disciolto nell’oceano (CO2, HCO3-, CO3 2-) è governata dal pH. L’acidificazione dell’oceano mediante l’anidride carbonica antropogenica sta causando una diminuzione delle concentrazioni di carbonato, gli atomi di carbonio delle molecole di carbonato si spostano invece nelle molecole di CO2 pertanto sta diventando dura per gli organismi sintetizzare le loro strutture di carbonato di calcio.
In questo modo Emiliania huxleyi è destinata alla pattumiera della storia?
Si estinguerà in futuro a causa dell’elevata CO2 dell’effetto serra? La risposta è che ciò potrebbe succedere ma non si sa ancora con certezza. Malgrado il pessimismo generale c’è tuttavia un raggio di speranza. Molti milioni di anni fa, nel Cretaceo (~145 a 65 milioni di anni fa) c’erano elevati livelli climatici di anidride carbonica paragonabili all’intensa CO2 dell’effetto serra che stiamo generando oggi. I Coccolitofori sono sopravvissuti (anche se Ehux non si era ancora evoluta) anche al caldo, ad elevata CO2 climatico, malgrado le basse concentrazioni dello ione bicarbonato. La presenza dei Coccolitofori in quel periodo è testimoniata dagli abbondanti coccoliti presenti nei sedimenti marini formatosi a quei tempi e dalla presenza di carbonato di calcio solido nelle profondità dell’oceano come lo è oggi. Ciò suggerisce che l’oceano era probabilmente sovrassaturo rispetto al carbonato di calcio come lo è oggi, malgrado l’alto contenuto di CO2. Ehux, come gli altri Coccolitofori, potranno trovarsi in difficoltà in futuro se l’oceano diventerà più acido a causa dell’elevata diffusione della CO2 dei combustibili fossili attraverso la superficie del mare. Ma si spera in ricerche future che dimostrino che tale preoccupazione è infondata in quanto sarebbe effettivamente una vergogna perdere questo favoloso fenomeno naturale.
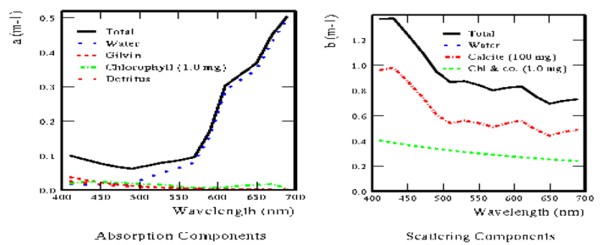
Fig.6. Componenti che contribuiscono all’assorbimento e alla riflessione nell’Atlantico a metà estate durante un bloom di Ehux. Gli effetti di clorofilla e calcite sono proporzionali alla quantità di sostanze nell’acqua.
Questo articolo è protetto da Copyright © e non può essere riprodotto e diffuso tramite nessun mezzo elettronico o cartaceo senza esplicita autorizzazione scritta da parte dello staff di BiologiaMarina.eu.
Ideazione: Pierfederici Giovanni - Progetto: Pierfederici Giovanni, Castronuovo Motta Nicola.
Prima Pubblicazione 31 Lug 2006 - Le immagini dei Collaboratori detentori del Copyright © sono riproducibili solo dietro specifica autorizzazione.
Si dichiara, ai sensi della legge del 7 Marzo 2001 n. 62 che questo sito non rientra nella categoria di "Informazione periodica" in quanto viene aggiornato ad intervalli non regolari
XHTML 1.0 Transitional – CSS
